
 English
English
Tin tức
Bổ Sung Các Chủng Bacillus Subtilis Trong Nước Nuôi Giúp Cải Thiện Chất Lượng Nước, Tăng Năng Suất Tăng Trưởng, Phản Ứng Miễn Dịch Và Khả Năng Chống Lại Vi Khuẩn Vibrio Harveyi Ở Tôm Thẻ Chân Trắng Litopenaeus Vannamei
Tóm tắt
Trong nghiên cứu này, huyền phù tế bào sinh dưỡng của hai chủng Bacillus subtilis, L10 và G1 với tỷ lệ bằng nhau, nhưng khác liều lượng, lần lượt là 105 (BM5) và 108 (BM8) CFU/ml đượ c sử dụng trong nước nuôi tôm thẻ chân trắng Litopenaeus vannamei trong thời gian 8 tuần. Cả hai chủng lợi khuẩn đều cho thấy nồng độ của các ion amoniac, nitrit và nitrat giảm đáng kể trong điều kiện phòng thí nghiệm và cả trên sinh vật sống. Các chỉ số về trọng lượng cuối cùng, tăng trọng, tốc độ tăng trưởng cụ thể (SGR), hệ số chuyển đổi thức ăn (FCR) và hoạt tính của các enzym tiêu hóa ở các nhóm BM5 và BM8 lớn hơn đáng kể so với nhóm đối chứng. Trong đó, nhóm BM8 và nhóm đối chứng nhận thấy sự khác biệt đáng kể về tỷ lệ sống. 8 tuần sau khi bắt đầu thử nghiệm, tôm được cảm nhiễm với vi khuẩn Vibrio harveyi. Kết quả phân tích thống kê cho thấy có sự khác biệt đáng kể về tỷ lệ sống của tôm giữa nhóm sử dụng probiotic và nhóm đối chứng. Tỷ lệ chết tích lũy của nhóm đối chứng là 80%, trong khi tỷ lệ chết tích lũy của nhóm sử dụng probiotic là 36,7% đối với nhóm BM8 và 50% đối với nhóm BM5. Sau đó, xét nghiệm PCR được sử dụng để xác định mức mRNA của propnoloxidase (proPO), peroxinectin (PE), lipopolysaccharide- và protein liên kết β-1,3-glucan (LGBP) và protein huyết thanh (SP). Sự biểu hiện của tất cả các gen liên quan đến miễn dịch trong nghiên cứu được điều chỉnh tăng đáng kể ở nhóm BM5 so với BM8 và nhóm đối chứng. Những kết quả này cho thấy rằng việc sử dụng các chủng B. subtilis trong nước nuôi mang lại những hiệu quả tích cực trong nuôi tôm, bao gồm các chỉ số về chất lượng nước, hiệu suất tăng trưởng, hoạt động của các enzym tiêu hóa, phản ứng miễn dịch và khả năng kháng bệnh.
Từ khóa: Nuôi trồng thủy sản, Litopenaeus vannamei, Bacillus subtilis, Vibriosis, Phản ứng miễn dịch
1/ Giới thiệu
Ngành nuôi tôm đã nhanh chóng được mở rộng nhờ vào sự phát triển của các hệ thống thâm canh trong nuôi trồng thủy sản; tuy nhiên, ngành công nghiệp này bị đe dọa bởi các tác nhân gây bệnh, chủ yếu là các mầm bệnh cơ hội như vi khuẩn Vibrio.
Việc sử dụng probiotic (chế phẩm sinh học – CPSH) đã được khuyến cáo thay cho việc sử dụng thuốc kháng sinh do sự xuất hiện của các vi khuẩn kháng kháng sinh. Các khía cạnh khác nhau của CPSH trong nuôi tôm đã được nghiên cứu và thu được nhiều kết quả đầy hứa hẹn. Ví dụ, việc bổ sung Pediococcus acidilactici vào chế độ ăn của tôm Litopenaeus stylirostris đã cải thiện tỷ lệ sống của chúng.
Các nghiên cứu trước đây cũng đã chứng minh rằng các chủng Bacillus subtilis có thể được sử dụng để làm CPSH tiềm năng trong nuôi tôm. Trên thực tế, Liu và cộng sự (2009) đã chứng minh rằng tôm thẻ chân trắng Litopenaeus vannamei được cho ăn với chế độ ăn bổ sung B. subtilis E20 có hiệu suất tăng trưởng tốt hơn so với nhóm đối chứng. Sau đó, Liu và cộng sự (2010) cho thấy rằng việc áp dụng loài này làm chất bổ sung trong nước giúp cải thiện tỷ lệ sống và phản ứng miễn dịch ở ấu trùng tôm thẻ chân trắng. Kết quả tương tự cũng đã được báo cáo bởi Shen và cộng sự (2010), rằng tôm thẻ chân trắng được nuôi với chế độ ăn bổ sung B. subtilis có hiệu suất tăng trưởng, phản ứng miễn dịch và hoạt động chống oxy hóa tốt hơn so với nhóm đối chứng.
Điều quan trọng cần đề cập là các cơ chế bảo vệ của tôm phần lớn phụ thuộc vào hệ thống miễn dịch bẩm sinh, và trên thực tế chúng không có khả năng phản ứng với các loại vắc xin cụ thể. Do đó, việc sử dụng B. subtilis như một CPSH tiềm năng có thể bù đắp cho những hạn chế của vắc xin trong nuôi tôm. Trong các nghiên cứu trước đây, chúng tôi đã báo cáo rằng tôm thẻ chân trắng được cho ăn chế độ ăn có chứa B. subtilis L10 và G1 có biểu hiện gen miễn dịch cao hơn so với nhóm đối chứng. Ngoài ra, tôm được bổ sung CPSH cũng cho thấy tỷ lệ sống cao hơn so với nhóm đối chứng sau khi được cảm nhiễm Vibrio harveyi.
Điều kiện môi trường và tình trạng nước nuôi là những mối quan tâm lớn trong nuôi tôm. Tình trạng nước nuôi bị ô nhiễm kéo dài có thể do chất lượng nước kém hoặc tỷ lệ thay nước thấp. Xem xét các điều kiện nuôi, các nhà nghiên cứu đã nhận thấy rằng vi khuẩn có đặc tính probiotic có thể hữu ích về mặt quản lý sức khỏe tôm. Tuy nhiên, lại có rất ít thông tin về tác dụng của chúng trong việc xử lý môi trường. Các nghiên cứu trước đây đã chứng minh rằng một số loài B. subtilis có khả năng tạo ra nhiều loại enzym ngoại bào và peptit kháng khuẩn. Việc tiết ra các chất này không chỉ giúp kiểm soát vi khuẩn gây bệnh mà còn giúp cải thiện chất lượng môi trường nước.
Một CPSH tiềm năng cần được đánh giá kỹ lưỡng về đặc tính và độ an toàn trước khi được thương mại hóa. Trước đây, chúng tôi đã báo cáo về việc phân lập, xác định, đặc tính và độ an toàn của 2 CPSH B. subtilis L10 và G1 đối với khả năng kháng với hai loài vi khuẩn phổ biến trên tôm là V. harveyi và Vibrio parahaemolyticus. Ngoài ra, gần đây chúng tôi đã đạt được những kết quả đầy hứa hẹn đối với việc bổ sung 2 CPSH này vào chế độ ăn của tôm thẻ chân trắng. Do đó, mục tiêu của nghiên cứu hiện tại là điều tra tác động của việc sử dụng CPSH B. subtilis L10 và G1 ở hai liều lượng khác nhau, trong nước nuôi để kiểm tra chất lượng nước, năng suất tăng trưởng, hoạt động của các enzym tiêu hóa, biểu hiện gen miễn dịch và khả năng kháng bệnh ở tôm thẻ chân trắng.
2/ Nguyên liệu và phương pháp
2.1. Chủng vi khuẩn
Trước đây, Bacillus subtilis L10 và G1 đã được phân lập và xác định từ dưa chua lên men, chúng được sử dụng như một CPSH tiềm năng. Vi khuẩn gây bệnh V. harveyi ATCC 14126 đã được sử dụng để cảm nhiễm trên tôm. Tất cả các chủng đều được giữ ở nhiệt độ 20 độ C trong môi trường Luria-Bertani Broth (LB; Difco) chứa 15% glycerol (v/v) cho đến khi sử dụng.
2.2. Thử nghiệm về nồng độ ion trong phòng thí nghiệm
Ảnh hưởng của hai chủng B. subtilis đến nồng độ ion được xác định bằng cách sử dụng nước ao tổng hợp (theo Lalloo và cộng sự, 2007). Việc chuẩn bị nước ao được thực hiện bằng cách sử dụng 0,0085% m/v KNO3, 0,006% m/v NaNO2, 0,0093% m/v (NH4)2SO4, 1% m/v NaCl, 0,1 m/v chiết xuất nấm men và 0,1% m/v glucose, với pH được điều chỉnh đến 7,0 và được khử trùng qua bộ lọc vô trùng 0,22 mm.
Mỗi chủng với 5 ml (đã được nuôi qua đêm) được thêm vào bể Erlenmeyer 1000 ml chứa 500 ml nước ao tổng hợp. Sau đó, các bình được ủ với nhiệt độ 30 độ C trên máy lắc quay ở tốc độ 150 vòng/phút. Mỗi bình được lấy mẫu vô trùng trước khi cấy ở 2 giờ, 8 giờ, 16 giờ và 24 giờ. Nồng độ nitrit, nitrat và amoniac được đánh giá quang phổ bằng cách sử dụng bộ HACH theo hướng dẫn của nhà sản xuất (HACH Company; Loveland, CO, USA). Nồng độ ion giảm được biểu thị bằng mg/L/giờ. Thử nghiệm cũng bao gồm một nhóm đối chứng không bổ sung CPSH. Sự phát triển của cả hai chủng trong nước nuôi tổng hợp cũng được theo dõi thông qua quang phổ kế bằng cách đo mật độ quang ở 600 nm (OD600) sau 0 giờ, 2 giờ, 8 giờ, 16 giờ và 24 giờ. Tất cả các thử nghiệm được lặp lại 3 lần.
2.3. Việc chuẩn bị chế phẩm sinh học
Hai chủng B. subtilis được nuôi trong môi trường LB sử dụng máy lắc ủ nhiệt ở 30 độ C trong 48 giờ. Sau đó, các vi khuẩn được ly tâm ở 3000 g trong 10 phút ở 4 độ C. Sau khi loại bỏ phần nổi phía trên, các vi khuẩn dạng viên được thu lại và rửa ba lần trong dung dịch muối vô trùng (NSS, 0,9% NaCl). Mật độ của huyền phù tế bào được tính toán bằng máy quang phổ ở bước sóng 600 nm và cũng tương quan với các đơn vị hình thành khuẩn lạc (CFU) bằng kỹ thuật trải đĩa. Các huyền phù tế bào sinh dưỡng này được giữ ở nhiệt độ 4 độ C, sau đó lập tức được đưa đến trại sản xuất tôm giống và bổ sung vào các bể nuôi.
2.4. Tôm và điều kiện thí nghiệm
Tôm giống khỏe mạnh được cung cấp bởi Trạm Nghiên cứu Khoa học Biển và Trạm thuộc lĩnh vực Sinh học, UPM, Port Dickson, Malaysia. Các thí nghiệm được tiến hành tại cùng một địa điểm. Những con tôm không bị nhiễm bệnh hoặc không có mầm bệnh được kiểm tra bằng các kỹ thuật phân tử tiêu chuẩn. Tôm được thuần 1 tuần trong các bể nuôi trước khi bắt đầu thử nghiệm. Sau thời gian thuần, trọng lượng trung bình của tôm là 0,67 g, và tôm được chia ngẫu nhiên vào 6 bể 500 lít, mỗi bể chứa 100 con. Tất cả tôm đều được cho ăn theo chế độ ăn thương mại (BLANCA, Malaysia) 3 lần/ngày (8 giờ sáng, 4 giờ chiều và 12 giờ đêm) ở mức 5% trọng lượng cơ thể. Các đơn vị thí nghiệm được duy trì trong điều kiện sục khí liên tục (5 ± 0,5% oxy hòa tan), với mức thay nước 50% 2 lần/tuần, nhiệt độ môi trường là 28 ± 1 độ C và pH là 7,3 – 8,2. Nhiệt độ, oxy hòa tan và pH được đo bằng máy quang phổ YSI (Yellow Spring Inc.). Các thông số hóa học trong nước như amoniac, nitrit và nitrat được đo quang phổ mỗi tuần một lần bằng bộ dụng cụ HACH như đã đề cập ở trên.
Huyền phù tế bào sinh dưỡng của các chủng B. subtilis L10 và G1 với tỷ lệ bằng nhau, được thêm vào nước nuôi để tạo ra nồng độ cuối cùng khoảng 105 CFU/ml đối với BM5 và 108 CFU/ml đối với BM8. Nồng độ thấp hơn được chọn dựa trên các nghiên cứu trước đây, nhưng nồng độ cao hơn được sử dụng bởi vì nó được cho là có hiệu quả hơn. Nhóm thứ ba không bổ sung CPSH là nhóm đối chứng. Tất cả các thí nghiệm được lặp lại trong 8 tuần và CPSH được bổ sung vào nước nuôi 2 lần một tuần.
Kết thúc thí nghiệm, các kết quả về trọng lượng cuối cùng, tỷ lệ sống, tăng trọng, hệ số chuyển đổi thức ăn (FCR), và tốc độ tăng trưởng cụ thể (SGR) giữa các nghiệm thức được tính toán dựa trên các phương pháp đã mô tả ở trên. 5 con tôm được thu thập ngẫu nhiên vào tuần 0, tuần 4 và tuần 8 sau mỗi lần lặp lại của thử nghiệm để ước tính số lượng Bacillus sp. và Vibrio sp. trong đường tiêu hóa (GI) của tôm. Thử nghiệm cũng thu thập 5 con tôm vào cuối tuần thứ 8 để kiểm tra hoạt tính enzym. Ống tiêu hóa của mỗi con tôm được cắt ra trong điều kiện vô trùng, sau đó được gói ngay lập tức và ngâm trong nitơ lỏng. Các mẫu được giữ ở nhiệt độ -80 độ C cho đến khi được phân tích. Tương tự, 2 con tôm từ mỗi lần lặp lại của thử nghiệm cũng được thu thập ngẫu nhiên để xác định mức độ biểu hiện của các gen liên quan đến miễn dịch, như propnoloxidase (proPO), peroxinectin (PE), lipopolysaccharide- và protein liên kết β -1,3-glucan (LGBP) và protein serine (SP).
Để ước tính số lượng của Bacillus sp. và Vibrio sp. trong đường tiêu hóa của tôm và nước nuôi, môi trường nuôi cấy chọn lọc đã được sử dụng như đã mô tả trước đó. Tôm được mổ bằng cách sử dụng một chiếc kéo phẫu thuật vô trùng, đường tiêu hóa được lấy ra, và đồng nhất trong thiết bị đồng nhất vô trùng với PBS. Để chắc chắn rằng có sự xâm nhập của B. subtilis trong đường tiêu hóa của tôm, vào cuối tuần thứ 8, DNA của 10 khuẩn lạc ngẫu nhiên từ các đĩa với số lượng khuẩn lạc có thể chấp nhận được (30 – 300 CFU/đĩa) đã được tách chiết và xác định bằng xét nghiệm PCR theo các điều kiện được mô tả.
2.5. Phân tích hoạt động của enzym
Dịch chiết thô của đường tiêu hóa được sử dụng để định lượng hoạt tính enzym tiêu hóa của tôm trong các nghiệm thức. Toàn bộ đường tiêu hóa của một hoặc hai con tôm được sẽ được mổ, thu lại, cân và đồng nhất với nước khử ion lạnh (1:10). Sau đó, chất đồng nhất sẽ được ly tâm với mức 5000 g trong 20 phút ở 4 độ C. Phần nổi phía trên được tách ra một cách cẩn thận và cho qua màng lọc với kích thước lỗ rỗng là 0,45 mm (Sartorius, Đức). Nước trong thử nghiệm được đựng trong 3 ống Eppendorf với thể tích 1,5 ml và giữ ở nhiệt độ 20 độ C để phân tích các enzym khác nhau.
Protein toàn phần được định lượng bằng cách sử dụng albumin huyết thanh bò theo tiêu chuẩn của Bradford. Tổng hoạt tính của protease được kiểm tra bằng cách sử dụng casein làm chất nền phản ứng với thuốc thử Folin. Đường chuẩn của độ hấp thụ ở bước sóng 440 nm được chuẩn bị bằng cách sử dụng tyrosine làm chất chuẩn. Một đơn vị hoạt động của protease được xác định là số lượng micromoles của tyrosine được giải phóng mỗi phút trên mỗi mg protein ở 37oC. Tổng hoạt tính amylase được xác định theo Rick và Stegbauer, sử dụng 1% tinh bột hòa tan làm chất nền phản ứng với axit 3,5-dinitrosalicylic. Đường chuẩn của độ hấp thụ ở bước sóng 550 nm được chuẩn bị bằng cách sử dụng dung dịch maltose chuẩn. Một đơn vị hoạt động của amylase được xác định là số lượng micromoles của maltose được giải phóng mỗi phút trên mỗi mg protein ở 37oC.
2.6. Thực nghiệm cảm nhiễm
Sau 8 tuần kể từ khi bổ sung CPSH vào nước nuôi, thử nghiệm đã tiến hành cho tôm cảm nhiễm với vi khuẩn gây bệnh V. harveyi ATCC 14126. Chủng vi khuẩn này được nuôi qua đêm trong môi trường LB, nồng độ được điều chỉnh đến 107 CFU/ml bằng cách sử dụng NSS như đã đề cập ở trên. 30 con tôm ở giai đoạn lột xác được thu thập từ các nhóm của thực nghiệm và cả nhóm đối chứng, sau đó tiêm 20 µl huyền phù vi khuẩn vào đoạn bụng thứ ba. Ngay sau khi tiêm, tôm được đưa vào bể 20 lít, mỗi bể 10 con. Thí nghiệm được thực hiện 3 lần và sử dụng nước từ các bể trước đó để giảm stress cho tôm. Nhóm tôm trước đó chưa được bổ sung B. subtilis được tiêm NNS, gọi là nhóm đối chứng tích cực (NB). Trong quá trình cảm nhiễm, tôm được cho ăn 3 lần mỗi ngày (8 giờ sáng, 4 giờ chiều và 12 giờ đêm). Tỷ lệ chết được theo dõi hàng ngày cho đến ngày thứ 10.
2.7. Biểu hiện mRNA của các gen liên quan đến miễn dịch
Sự biểu hiện của các gen liên quan đến miễn dịch ở tôm do chủng B. subtilis tác động trong quá trình cảm nhiễm với V. harveyi được xác định bằng xét nghiệm PCR (RT-qPCR). Hai con tôm từ mỗi lần lặp lại của thử nghiệm được lấy ngẫu nhiên để tách chiết RNA. Vì hầu như không thể thu thập được các tế bào huyết cầu từ tôm (3.0 – 4.0 g), nên tôm được làm đông bằng cách sử dụng một lượng lớn nitơ lỏng và được đồng nhất bằng cối và chày. 100 mg chất đồng nhất được sử dụng để chiết xuất và tinh chế RNA bằng phương pháp guanidinium thiocyanate. Phiên mã ngược QuantiTect (Qiagen) chứa oligo- (dT) được sử dụng để tổng hợp cDNA. Các bước chuẩn bị đều tuân theo khuyến nghị của nhà sản xuất để tối đa hóa quá trình tổng hợp cDNA. Xét nghiệm PCR được thực hiện với các đoạn mồi sau để xác định mức độ biểu hiện của các gen liên quan đến miễn dịch: đối với ProPO, thuận (5’-GCC TTG GCA ACG CTT TCA-3’) và ngược (5’-GC GCA TCA GTT CAG TTT GT-3’ ); đối với PE, thuận (5’-TGG ACC TCG CGG GAG AT-3’) và nghịch (5’-GAC CGA TAG CCA CCA TGC TT-3’); đối với LGBP, thuận (5’-CAT GTC CAA CTT CGC TTT CAGA-3’) và nghịch (5’-ATC ACC GCG TGG CAT CTT-3’); đối với SP, thuận (5’-CGT CGT TAG GTT AAG TGC GTT CT-3’) và nghịch (5’-TTT CAG CGC ATT AAG ACG TGTT-3R’). Mức độ biểu hiện được chuẩn hóa bằng cách sử dụng bactin (thuận 5’-GAG CAA CAC GGA GTT CGT TGT-3’ và nghịch 5’- CAT CAC CAA CTG GGA CGA CAT GGA-3’) làm gen giữ nhà, tuân theo các điều kiện đã được mô tả.
2.8. Phân tích thống kê
Dữ liệu về các thông số tăng trưởng, hoạt tính của các enzym tiêu hóa, phân tích vi khuẩn, phản ứng miễn dịch và khả năng kháng mầm bệnh giữa các nghiệm thức được phân tích bằng cách sử dụng ANOVA và trắc nghiệm đa đoạn Duncan được sử dụng để xác định sự thay đổi đáng kể (P <0,05). Tất cả các phân tích thống kê được thực hiện bằng SPSS, phiên bản 15 (SPSS Inc, Chicago, IL, USA).
3/ Kết quả
3.1. Thử nghiệm về nồng độ ion trong phòng thí nghiệm
Nghiên cứu sử dụng nước tổng hợp để đánh giá khả năng khử các ion amoniac, nitrit và nitrat của B. subtilis L10 và G1 trong 24 giờ (Bảng 1). Cả hai chủng đều cho thấy khả năng giảm nồng độ amoniac và nitrit tương tự nhau với tỷ lệ lần lượt là 0,044 và 0,53 mg/l/giờ. Khả năng khử nitrat đối với L10 là 2,43 mg/l/giờ và G1 là 2,79 mg/l/giờ. Ngoài ra, cả hai chủng đều cho thấy mức tăng trưởng giống nhau trong nước ao tổng hợp. Mức tăng trưởng cao nhất được ghi nhận ở 16 giờ đối với cả hai chủng và trong 24 giờ quan sát thấy mật độ của quần thể vi khuẩn giảm (Bảng 1).
3.2. Thử nghiệm về chất lượng nước trên cơ thể sinh vật
Bắt đầu từ tuần thứ 5 cho đến khi kết thúc thí nghiệm, nghiên cứu nhận thấy có sự khác biệt đáng kể (P <0,05) về nồng độ amoniac giữa các nhóm bổ sung CPSH và nhóm đối chứng (Hình 1). Sự khác biệt này dao động từ 0,22 ± 0,08 – 0,78 ± 0,2 mg/l và 0,17 ± 0,2 – 0,66 ± 0,35 mg/l ở nhóm BM8 và BM5, trong khi nhóm đối chứng đạt giá trị từ 1,04 ± 0,2 mg/l đến nồng độ cao nhất là 1,58 ± 0,45 mg/l khi kết thúc thí nghiệm. Không có sự khác biệt đáng kể (P> 0,05) về nồng độ nitrit giữa tất cả các nhóm thí nghiệm từ khi bắt đầu thí nghiệm đến tuần thứ 5. Tuy nhiên, từ tuần thứ 6 đến khi kết thúc thí nghiệm, có sự khác biệt đáng kể (P <0,05) về nồng độ nitrit giữa nhóm BM8, so với nhóm BM5 và nhóm đối chứng. Nồng độ nitrit giảm mạnh ở nhóm BM8, dao động từ 0,007 ± 0008 – 0,0007 ± 0,0015 mg/l, nhưng lại tăng nhẹ ở nhóm BM5 và nhóm chứng, dao động lần lượt là 0,0097 ± 0,0017 – 0,012 ± 0,001 mg/l và 0,010 ± 0,002 – 0,125 ± 0,001 mg/l (Hình 1). Sự khác biệt về nồng độ nitrat (P <0,05) chỉ quan sát thấy ở tuần thứ 7 và thứ 8. Nhóm BM8 có nồng độ cao nhất (P <0,05) từ 3,72 ± 0,3 – 4 ± 0,18 mg/l. Tiếp theo là nhóm chứng với 2,72 ± 0,59 – 3,07 ± 0,35 mg/l và cuối cùng là nhóm BM5 với 1,1 ± 0,29 – 1,55 ± 0,31 mg/l) (Hình 1).
Bảng 1: Phản ứng của B. subtilis L10 và G1 đối với quá trình khử ion trong nước biển tổng hợp.
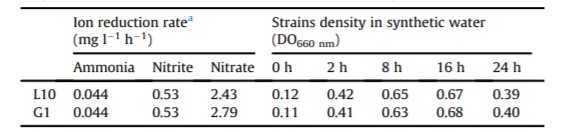
a: Tốc độ khử ion được tính dựa trên thời gian ủ là 24 giờ.
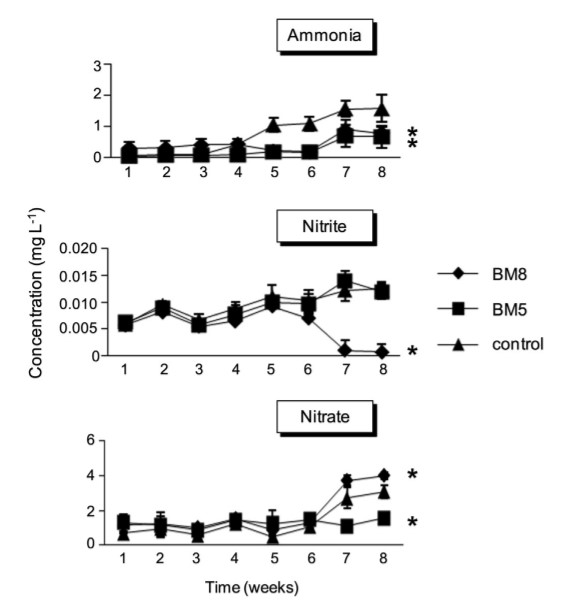
Hình 1. Sự thay đổi nồng độ amoniac, nitrit và nitrat dư trong nước nuôi của tôm thẻ chân trắng đối với (♦) BM8 (B. subtilis L10 và G1, 108 CFU/ml); (■) BM5 (B. subtilis L10 và G1, 105 CFU/ml); và (▲) nhóm đối chứng. Dấu hoa thị biểu thị ý nghĩa thống kê, P <0,05, so với nhóm đối chứng.
3.3. Các thông số tăng trưởng
Khi bắt đầu thí nghiệm, không có sự khác biệt đáng kể (P> 0,05) về trọng lượng giữa nhóm được bổ sung CPSH và nhóm đối chứng. Tuy nhiên, nghiên cứu nhận thấy có sự khác biệt (P <0,05) về trọng lượng cuối cùng, tăng trọng, FCR và SGR của tôm giữa các nhóm được bổ sung CPSH và nhóm đối chứng vào cuối thí nghiệm (Bảng 2). Ngoài ra, không quan sát thấy sự khác biệt (P> 0,05) về tỷ lệ sống giữa nhóm BM8 và BM5 cũng như giữa nhóm BM5 và nhóm đối chứng; tuy nhiên, có sự khác biệt đáng kể (P <0,05) về tỷ lệ sống của những con tôm ở nhóm BM8 so với nhóm đối chứng (Bảng 2).
Bảng 2: Dữ liệu về hiệu suất tăng trưởng và tỷ lệ sống của tôm thẻ chân trắng giữa các nhóm được bổ sung CPSH và nhóm đối chứng
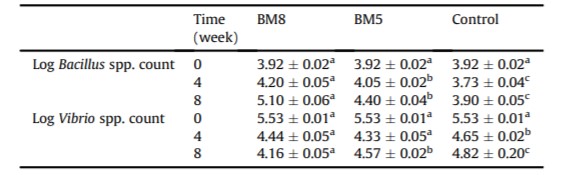
Bảng 3: Giá trị trung bình về số lượng Bacillus spp. và Vibrio spp. (CFU/g) trong đường tiêu hóa của tôm thẻ chân trắng giữa các nhóm được bổ sung CPSH và nhóm đối chứng trong 55 ngày.
3.4. Phân tích vi khuẩn
Số lượng Bacillus spp. và Vibrio spp. trong đường tiêu hóa tôm và nước nuôi ở tuần 0, tuần 4 và tuần 8 được trình bày trong Bảng 3 và 4. Có sự khác biệt đáng kể (P <0,05) về số lượng Bacillus spp. và Vibrio spp. trong đường tiêu hóa của tôm giữa các nhóm được bổ sung CPSH và nhóm đối chứng. Vào cuối thí nghiệm, số lượng của Bacillus spp. trong đường tiêu hóa của tôm ở nhóm BM8 và BM5 tăng lên đáng kể, lần lượt là 3,92 ± 0,02 – 5,1 ± 0,06 và 4,4 ± 0,04 log CFU/g. Ngược lại, số lượng Bacillus spp trong đường tiêu hóa của tôm ở nhóm đối chứng được nhận thấy là tương đối thấp (Bảng 3). Để xác nhận đặc điểm phân loại của chúng ở cấp độ loài, các khuẩn lạc màu vàng được đếm trên thạch MYP là Bacillus spp. được chọn ngẫu nhiên (n = 10) và được khuếch đại DNA bằng cách sử dụng các đoạn mồi cụ thể. Các phân tích vi khuẩn và phân tử cho thấy B. subtilis L10 và G1 có khả năng xâm nhập vào đường tiêu hóa của tôm. Hơn nữa, số lượng Vibrio spp. trong đường tiêu hóa của tôm ở nhóm BM8 và BM5 giảm đáng kể vào tuần thứ 4 và thứ 8, so với nhóm đối chứng (Bảng 3).
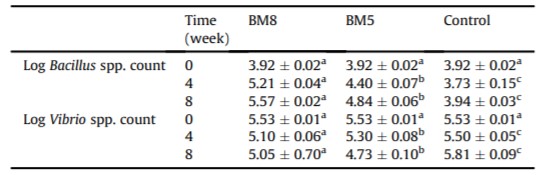
Số lượng Bacillus spp. và Vibrio spp. trong nước nuôi giữa các nhóm được bổ sung CPSH và nhóm đối chứng là khác nhau đáng kể trong các giai đoạn thí nghiệm. Số lượng Bacillus spp. trong nước nuôi ở nhóm BM8 và BM5 cao hơn đáng kể (P <0,05) so với nhóm đối chứng ở tuần thứ 4 và thứ 8 (Bảng 4). Số lượng Vibrio spp. ở nhóm chứng cao hơn đáng kể (P <0,05) so với nhóm BM8 và BM5 ở tuần thứ 4 và thứ 8 (Bảng 4).
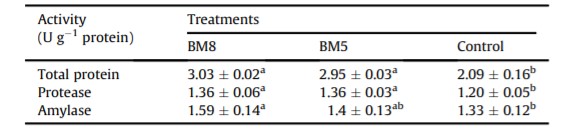
Protein toàn phần, protease và amylase trong đường tiêu hóa của tôm được đánh giá và so sánh thống kê trong Bảng 5. Có sự khác biệt đáng kể về protein toàn phần và hoạt động của protease trong đường tiêu hóa của tôm ở nhóm BM8 và BM5 so với nhóm đối chứng (Bảng 5). Hoạt tính của amylase trong đường tiêu hóa của tôm ở nhóm BM8 (1,59 ± 0,14 U/g protein) được nhận thấy là cao hơn đáng kể so với trong nhóm đối chứng. Tuy nhiên, không có sự khác biệt nào được nhận thấy về hoạt tính amylase giữa nhóm BM5 (1,4 ± 0,13 U/g protein) và nhóm đối chứng (1,33 ± 0,12 U/g protein). Hoạt tính của protein toàn phần và protease (P <0,05) trong đường tiêu hóa của tôm ở nhóm BM8 và BM5 cao hơn so với nhóm đối chứng.
Bảng 4: Giá trị trung bình của Bacillus spp. và Vibrio spp. (CFU/ml) trong nước nuôi của tôm thẻ chân trắng giữa các nhóm được bổ sung CPSH và nhóm đối chứng trong 55 ngày.
Bảng 5: Hoạt tính của enzym tiêu hóa ở tôm thẻ chân trắng giữa các nhóm được bổ sung CPSH và nhóm đối chứng trong 55 ngày.
3.6. Tỷ lệ chết tích lũy và biểu hiện gen liên quan đến miễn dịch
Sau 8 tuần thử nghiệm, tất cả các nhóm được cảm nhiễm với V. harveyi (Hình 2) với liều lượng 106 CFU/con. Kết quả cho thấy có sự khác biệt đáng kể (P <0,05) về tỷ lệ chết tích lũy ở nhóm BM8 (36,67 ± 5,77%) và BM5 (50 ± 10%), so với nhóm chứng (80 ± 10%). Không quan sát thấy tỷ lệ chết đối với tôm ở nhóm đối chứng sau khi được tiêm NSS.
Biểu hiện mRNA của bốn gen liên quan đến miễn dịch ở tất cả các nhóm trong thử nghiệm được đo sau 24 giờ cảm nhiễm bằng cách sử dụng các xét nghiệm PCR. Có sự khác biệt đáng kể về việc điều chỉnh tăng phiên mã proPO, PE, LGBP và SP ở tôm từ nhóm BM5, so với nhóm BM8 và nhóm đối chứng (Hình 2). Hơn nữa, biểu hiện LGBP cũng được điều chỉnh cao hơn đáng kể ở nhóm BM8 so với nhóm đối chứng, nhưng thấp hơn so với nhóm BM5 (Hình 3).
4/ Thảo luận
Các nghiên cứu trước đây đã chứng minh đặc tính của các CPSH thuộc chủng Bacillus ở một số sinh vật thủy sinh. Trong nghiên cứu hiện tại, hai chủng B. subtilis L10 và G1 cho thấy nồng độ của các ion amoniac, nitrit và nitrat trong điều kiện phòng thí nghiệm giảm đáng kể. Sau đó, việc bổ sung các chủng vi khuẩn này trong nước nuôi đã duy trì nồng độ của các ion amoniac, nitrit và nitrat trong phạm vi có thể chấp nhận được đối với tôm nuôi, đặc biệt là trong 3 tuần cuối của thí nghiệm. Mặc dù những phát hiện này phù hợp với kết quả của các nghiên cứu trước, nhưng Liu và cộng sự (2009) cho thấy rằng B. subtilis E20 không góp phần vào việc cải thiện chất lượng nước. Tuy nhiên, việc so sánh trực tiếp giữa các lợi khuẩn này rất khó vì chúng thường khác nhau về chức năng và có thể bị ảnh hưởng bởi các yếu tố khác nhau như di truyền, dinh dưỡng và môi trường.
Kết quả của nghiên cứu này cũng chứng minh rằng việc bổ sung các chủng B. subtilis trong nước nuôi có tác động tích cực đến trọng lượng cuối cùng, tăng trọng, SGR, FCR và tỷ lệ sống của tôm hơn so với nhóm đối chứng. Các nghiên cứu trước đây đã chỉ ra rằng việc sử dụng CPSH làm chất phụ gia trong nước có thể cải thiện hiệu suất tăng trưởng và tỷ lệ sống của tôm. Việc đường tiêu hóa của tôm tiết ra các enzym tiêu hóa có thể là một cách giải thích khác cho hiệu suất tăng trưởng tốt hơn, đồng thời nó cũng giúp tôm có sức khỏe và tỷ lệ sống cao hơn. Một số nghiên cứu đã chứng minh rằng các enzym của chủng Bacillus rất hiệu quả trong việc phân hủy nhiều loại carbohydrate, lipid và protein thành các đơn vị nhỏ hơn. Ngoài ra, việc sử dụng các chủng B. subtilis này cũng làm tăng hoạt tính của protein toàn phần, protease và amylase trong đường tiêu hóa của tôm. Do đó, sự hiện diện của những lợi khuẩn này trong đường tiêu hóa đã góp phần cải thiện khả năng tiêu hóa thức ăn, từ đó có thể dẫn đến tốc độ tăng trưởng hiệu quả hơn.
CPSH có tác dụng loại trừ cạnh tranh giúp cải thiện sự cân bằng hệ vi sinh vật trong đường ruột. Phương thức hoạt động khả thi này dẫn đến sự thay thế các vi khuẩn có lợi trong đường tiêu hóa, góp phần nâng cao hiệu suất tăng trưởng và tỷ lệ sống. Trong nghiên cứu hiện tại, nhóm được bổ sung các chủng B. subtilis cho thấy khả năng loại trừ cạnh tranh mạnh mẽ và sự xâm chiếm thành công trong đường tiêu hóa của tôm hơn so với nhóm đối chứng. Các chủng vi khuẩn này cũng tạo ra khả năng kháng V. harveyi cao hơn so với những con tôm không được bổ sung CPSH. Kết quả này tương tự với những kết quả được mô tả bởi các nhà nghiên cứu trước. Trên thực tế, Balcázar & Rojas-Luna (2007) đã chỉ ra rằng việc bổ sung B. subtilis UTM 126 vào chế độ ăn của tôm thẻ chân trắng có thể bảo vệ chúng chống lại bệnh vi khuẩn Vibrio. Ngoài ra, Zokaeifar và cộng sự (2012) cho thấy rằng việc bổ sung B. subtilis trong chế độ ăn làm giảm tỷ lệ chết của tôm sau khi cảm nhiễm với V. harveyi.
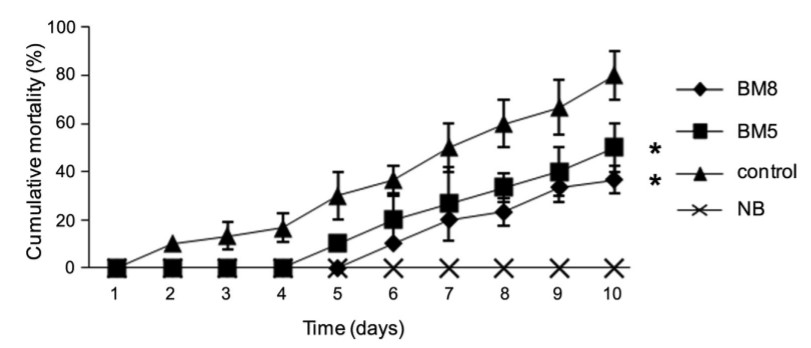
Hình 2. Tỷ lệ chết tích lũy (%) của tôm thẻ chân trắng 10 ngày sau khi cảm nhiễm với V. harveyi. (♦) BM8 (B. subtilis L10 và G1, 108 CFU/ml); (■) BM5 (B. subtilis L10 và G1, 105 CFU/ml); (×) NB (không bổ sung probiotic nhưng được tiêm NSS); và (▲) nhóm đối chứng (không bổ sung probiotic nhưng cảm nhiễm với vi khuẩn V. harveyi). Dấu hoa thị biểu thị ý nghĩa thống kê, P <0,05, so với nhóm đối chứng.
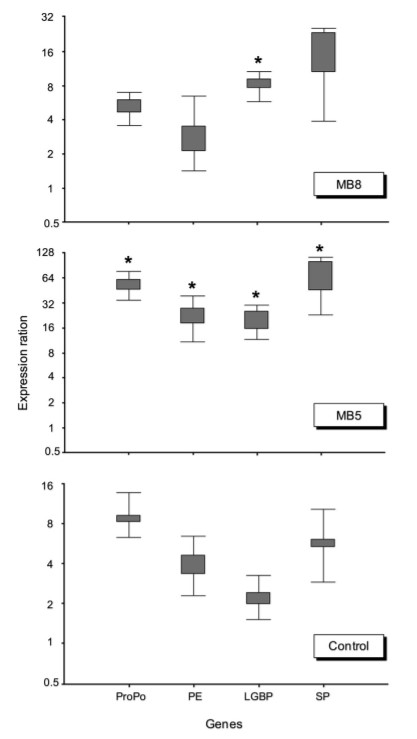
Hình 3. Biểu hiện mRNA của bốn gen liên quan đến miễn dịch của tôm thẻ chân trắng khi được bổ sung hoặc không bổ sung probiotic trong 55 ngày và khi cảm nhiễm với V. harveyi. Tất cả các phân tích được thực hiện bằng phần mềm REST. Các hộp đại diện cho khoảng biến thiên tứ phân vị, hoặc 50% giữa các giá trị. Râu đại diện cho các giá trị lớn nhất và nhỏ nhất. Dấu hoa thị biểu thị ý nghĩa thống kê, P <0,05, so với nhóm đối chứng.
Để đánh giá xem việc sử dụng CPSH có ảnh hưởng đến sự biểu hiện của các gen liên quan đến miễn dịch hay không, cả hai chủng B. subtilis đều được sử dụng ở hai liều lượng khác nhau, BM5 và BM8, trong nước nuôi và kết quả cho thấy có sự khác biệt đáng kể đối với sự điều hòa của proPO, PE (protein kết dính tế bào), LGBP (protein nhận biết) và SP (enzyme quan trọng liên quan đến hoạt động và kích hoạt PO) ở tôm của nhóm BM5, so với BM8 và nhóm đối chứng. Kết quả của nghiên cứu cũng chứng minh rằng biểu hiện LGBP ở nhóm BM8 cao hơn đáng kể so với nhóm chứng. Sự kích hoạt của hệ thống proPO là thông qua các phân tử nhận dạng trong dịch tương của động vật không xương sống, có thể nhận ra và phản ứng với những tác nhân xâm nhập thông qua lipopolysaccharides hoặc peptidoglycan từ vi khuẩn và β-1,3-glucans từ nấm. Sự liên kết của LGBP với lipopolysaccharides hoặc β -1,3-glucans đã được ghi nhận để kích hoạt hệ thống proPO của động vật giáp xác. Do đó, những phát hiện này chỉ ra rằng phản ứng miễn dịch chịu trách nhiệm một phần trong việc giúp kháng bệnh.
Mặc dù cả hai nghiệm thức BM5 và BM8 đều mang lại hiệu quả tích cực cho việc nuôi tôm, nhưng liều lượng tốt nhất để bổ sung nên là 108 CFU/ml (BM8). Điều này sẽ mang lại kết quả tốt về chất lượng nước, năng suất tăng trưởng, hoạt động của các enzym tiêu hóa, loại trừ cạnh tranh, phản ứng miễn dịch và kháng bệnh.
Theo Hadi Zokaeifar, Nahid Babaei, Che Roos Saad, Mohd Salleh Kamarudin, Kamaruzaman Sijam và Jose Luis Balcazar
Nguồn: http://dx.doi.org/10.1016/j.fsi.2013.10.007
Biên dịch: Huyền Thoại – Bình Minh Capital
Xem thêm:
- Nghiên Cứu Tầm Quan Trọng Của Vi Tảo Trong Ngành Nuôi Trồng Thủy Sản
- Sức Khỏe Của Hệ Vi Sinh Vật Đường Ruột Ở Tôm Post L. Vannamei Bị Ảnh Hưởng Sau Khi Giảm Độ Mặn
- Yếu Tố Ảnh Hưởng Lớn Nhất Đến Sự Đa Dạng Của Hệ Vi Sinh Vật Đường Ruột Ở Tôm Sú Là Thời Gian Chứ Không Phải Là Nguồn Protein