
 English
English
Kỹ Thuật Nuôi, Tin tức
Phần 2: Ảnh Hưởng Của Tình Trạng Nhịn Ăn Trong Thời Gian Ngắn Và Lột Xác Đến Hành Vi Kiếm Ăn Của Tôm Thẻ Chân Trắng (Litopenaeus vannamei)
3. Kết quả
3.1. Sự khác biệt về hành vi liên quan đến tình trạng nhịn ăn và lột xác
Các tác động chính đáng kể của tình trạng nhịn ăn (F5,173 = 4.350, p<0.01) và tình trạng lột xác (F5,173 = 4.776, p<0.001) đã được tìm thấy liên quan đến các hành vi trạng thái. Tuy nhiên, liên quan đến hành vi sự kiện, chỉ có tình trạng lột xác mới có tác dụng đáng kể trong mô hình (F2,99 = 3.199; p<0.05). Vì không có sự tương tác đáng kể nào giữa tình trạng nhịn ăn và lột xác được tìm thấy đối với các hành vi trạng thái hoặc sự kiện, các tác động chính của từng tình trạng được phân tích độc lập liên quan đến các hành vi được quan sát.
Nhịn ăn: Nhìn chung, một sự khác biệt đáng kể về thời gian thực hiện từng hành vi trạng thái đã được tìm thấy (F2,49 = 11.032, p<0.001) với sự tương tác đáng kể giữa thời gian nhịn ăn và các hành vi được thực hiện (F5,49 = 3.108, p < 0.01, Hình 2A). Tôm thuộc nhóm đối chứng dành nhiều thời gian không hoạt động và thực hiện các hành vi phát hiện hơn bất kỳ hành vi nào khác (F3,95=47,507, p<0,001). Tuy nhiên, với sự gia tăng thời kỳ nhịn ăn, sự khác biệt giữa các hành vi thụ động và chủ động đã biến mất (Hình 2A). Khi thời gian thực hiện cùng một hành vi được so sánh giữa các nghiệm thức, các hành vi bị ảnh hưởng bởi nghiệm thức nhịn ăn bao gồm không hoạt động (F2,63 = 4,092, p<0,05), phát hiện (F2,63 = 10,343, p< 0,001) và kiếm ăn (F2,63 = 6,312, p< 0,01). Phân tích post hoc của Tukey cho thấy sự khác biệt đáng kể giữa các nhóm đối chứng và +48 giờ nhịn ăn liên quan đến không hoạt động (p<0,05) và giữa nhóm đối chứng và các nhóm nhịn ăn khác để phát hiện (+24 giờ, p<0,01; +48 giờ, p<0,001) và kiếm ăn (+24 giờ, p <0,05; +48 giờ, p<0,01; Sung. 2A).
Một ảnh hưởng đáng kể của việc nhịn ăn cũng được tìm thấy trong việc thực hiện các hành vi sự kiện (F2,27 = 39,516, p<0,001; hình 2B). Sự khác biệt về số lần tôm thể hiện các hành vi này cũng được tìm thấy trong phạm vi đối chứng (F2,62 = 1,947, p<0,001), +24 giờ (F2,64 = 24,476, p<0,001) và +48 giờ (F2,63 = 30,528, p<0,001) các nhóm nhịn ăn, với số lần lau râu lớn hơn so với nhịp mắt và nhịp chân (Hình 2B). Tuy nhiên, không có sự khác biệt đáng kể nào được tìm thấy trong mỗi hành vi sự kiện theo thời gian nhịn ăn.
Tình trạng lột xác. Nói chung, một sự khác biệt đáng kể về thời gian thực hiện từng hành vi của trạng thái đã được tìm thấy (F2,177 = 19.240, p < 0.001). Một sự tương tác đáng kể giữa tình trạng và hành vi lột xác cũng đã được quan sát thấy (F5,177 = 4.269, p<0.01, Hình 2C) và có sự khác biệt về thời gian thực hiện các hành vi khác nhau cho mỗi nhóm trong số ba nhóm trạng thái lột xác (lột xác: F2,43 = 6.195, p<0.01; trước khi lột xác: F2,57 = 10.303, p<0.001; sau lột xác: F2,45 = 11.135, p<0.001). Tôm sau lột xác dành phần lớn thời gian để thực hiện các hành vi thụ động và thời gian kiếm ăn ít hơn đáng kể so với các hành vi còn lại (p<0,01 tất cả). Tuy nhiên, sự khác biệt này giữa hành vi thụ động và kiếm ăn không được nhìn thấy ở tôm trước khi lột xác hoặc liên moult, nơi những hành vi đó được phân bổ đều hơn trong thời gian ghi (Hình 2C). Khi mỗi hành vi được so sánh giữa tình trạng lột xác, chỉ có thời gian kiếm ăn là khác nhau đáng kể (F2,63 = 15,030, p<0,001), với tôm liên lột xác dành nhiều thời gian kiếm ăn hơn tôm trước khi lột xác (p<0,05) và tôm sau lột xác (p<0,001). Ngoài ra, tôm trước khi lột xác dành nhiều thời gian kiếm ăn hơn tôm sau khi lột xác (p<0,05) (Hình 2C).
Sự khác biệt đã được tìm thấy trong tần suất của các hành vi sự kiện (F2,121 = 50.223, p<0.001), với ảnh hưởng đáng kể đến tình trạng lột xác (F2,121 = 3.007, p<0.05; hình 2D). Liên lột xác (F2,28 = 28.972, p<0.001) và sau lột xác (F2,46 = 19.805, p<0.001) thực hiện số lần lau râu cao hơn so với nhịp mắt và nhịp tối đa, trong khi các cá thể trước khi lột xác (F2,46 = 12.540, p<0.001) thể hiện số lần nhịp mắt thấp hơn so với hai hành vi còn lại (Hình 2D). Hơn nữa, khi mỗi hành vi được so sánh giữa trạng thái lột xác, một sự khác biệt đáng kể đã được tìm thấy liên quan đến số lượng nhịp mắt (F2,63 = 6,250, p<0,01) và lau râu (F2,63 = 5,743, p<0,01; hình 2D). Các thử nghiệm của Tukey cho thấy tôm sau lột xác thực hiện số lần nhịp mắt cao hơn so với tôm trước khi lột xác (p<0,01) và liên lột xác (p<0,05). Tôm sau lột xác cũng thực hiện số lần lau râu cao hơn so với tôm trước khi lột xác (p<0,01) và liên lột xác (p<0,05) (Hình 2D).
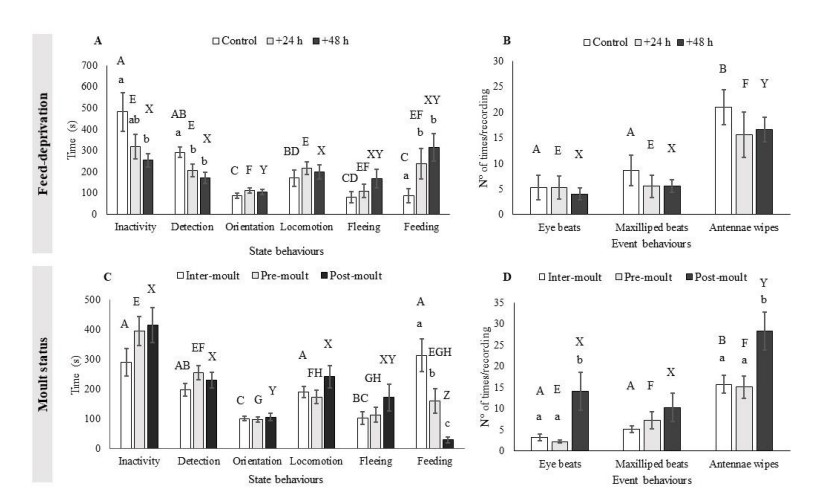
Hình 2. (Các) thời gian trung bình thực hiện từng hành vi trạng thái và hành vi sự kiện theo nhịn ăn (A, B; n = 8 tôm mỗi nghiệm thức nhịn ăn) và tình trạng lột xác (C, D; n = 24 con tôm mỗi lần lột xác vì mỗi con tôm trải qua cả ba giai đoạn lột xác, do đó các thanh đại diện cho các phép đo lặp đi lặp lại trên cùng một cá thể). Trình bày với các lỗi tiêu chuẩn. Sự khác biệt đáng kể (Tukey: p<0,05) về thời gian thực hiện các hành vi khác nhau của tôm trong cùng một nghiệm thức được biểu thị bằng chữ in hoa, trong đó các thanh chia sẻ chữ cái không khác nhau về mặt thống kê.
3.2. Mối tương quan giữa các hành vi được phân tích
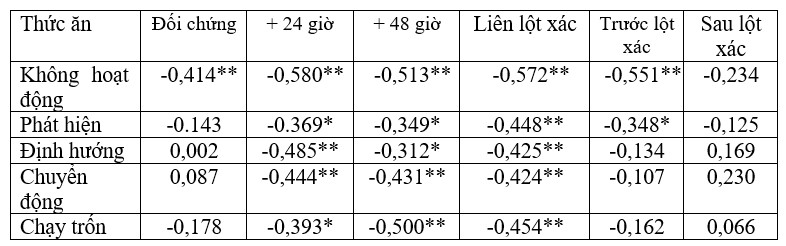
Quỹ thời gian tôm thay đổi theo cả thời kỳ nhịn ăn và tình trạng lột xác (Bảng 2). Có một xu hướng chung là hành vi kiếm ăn có mối tương quan tiêu cực với phần còn lại của các hành vi được đo lường. Tuy nhiên, đối với tình trạng nhịn ăn, chỉ có sự không hoạt động có mối tương quan tiêu cực đáng kể với hành vi kiếm ăn trong nhóm đối chứng (-0,414, p<0,001) trong khi tất cả các hành vi cho thấy mối tương quan tiêu cực đáng kể với việc kiếm ăn ở mức nhịn ăn +24 giờ và +48 giờ (Bảng 2A). Để lột xác, không có hành vi nào có mối tương quan tiêu cực đáng kể với việc kiếm ăn ở nhóm sau lột xác và trong nhóm trước khi lột xác, mối tương quan tiêu cực với việc kiếm ăn đạt được ý nghĩa với việc không hoạt động (-0,551, p<0,001) và phát hiện (-0,348, p<0,05). Tất cả các hành vi cho thấy mối tương quan tiêu cực đáng kể với việc kiếm ăn trong nhóm liên lột xác (Bảng 2B).
Bảng 2. Hệ số tương quan (r) giữa các hành vi khác nhau và việc cho ăn bằng cách sử dụng tất cả các quan sát video được thực hiện (120 quan sát hành vi) được xem xét liên quan đến (A) thời kỳ thiếu thức ăn (đối chứng, +24 giờ, +48 giờ (n = 8 con tôm mỗi nghiệm thức và 5 video quan sát trên mỗi con tôm = 40 video mỗi nghiệm thức)) và sau đó (B) tình trạng lột xác. Số lượng video của từng con tôm riêng lẻ không giống nhau cho mỗi giai đoạn lột xác do sự khác biệt về độ dài của từng giai đoạn, do đó có 52 quan sát hành vi của tôm liên lột xác, 44 con tôm trước khi lột xác và 24 con tôm sau lột xác). Các mối tương quan đáng kể được in đậm và được biểu thị là * p<0,05, ** p < 0,001. Các tế bào bóng mờ thể hiện mối tương quan tiêu cực với hành vi cho ăn.
3.3. Điểm hấp dẫn của thức ăn
Sự khác biệt đáng kể về điểm hấp dẫn thức ăn đã được tìm thấy liên quan đến nghiệm thức nhịn ăn (F2,63 = 3.817, p<0.05; hình 3A) và tình trạng lột xác (F2,63 = 4.374, p<0.05; hình 3B). Nhóm nhịn ăn +48 giờ và trạng thái liên lột xác có điểm hấp dẫn thức ăn cao nhất và khác biệt đáng kể lần lượt so với nhóm đối chứng và nhóm sau lột xác (kiểm nghiệm Tukey: p<0.05 cả hai so sánh).
Hơn nữa, thời gian cho từng con tôm ăn có thể được dự đoán mạnh mẽ bằng điểm hấp dẫn thức ăn (R2 = 0,615, p<0,001; hình 3C). Tôm có điểm hấp dẫn thức ăn cao nhất và do đó thời gian kiếm ăn lâu hơn chủ yếu đến từ nhóm nhịn ăn +48 giờ trong khi tôm có điểm hấp dẫn thức ăn thấp nhất là từ nhóm đối chứng. Tôm bị nhịn ăn trong +24 giờ được phân phối dọc theo phạm vi hấp dẫn thức ăn (Hình 3C).
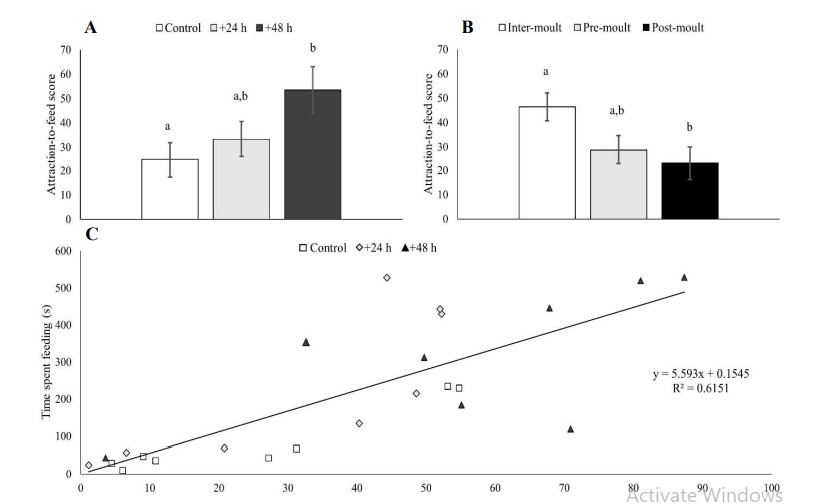
Hình 3. Điểm hấp thụ thức ăn (%) của tôm theo thời kỳ nhịn ăn (A; n = 8 con mỗi nghiệm thức nhịn ăn) và tình trạng lột xác (B; n = 24 con mỗi lần lột xác vì mỗi con tôm trải qua cả ba giai đoạn lột xác, do đó các thanh đại diện cho các phép đo lặp đi lặp lại trên cùng một cá thể). Trình bày với các lỗi tiêu chuẩn. Sự khác biệt đáng kể (Tukey: p<0,05; chữ thường) đã được tìm thấy theo thời gian thiếu thức ăn giữa đối chứng và nhịn ăn +48 giờ và giữa các cá thể liên và sau khi lột xác liên quan đến tình trạng lột xác. (C) Mối tương quan giữa điểm thu hút thức ăn của mỗi cá nhân và thời gian cho ăn. Các nhóm nhịn ăn được xác định bằng các ký hiệu; hình vuông = đối chứng, kim cương = +24 giờ nhịn ăn, hình tam giác = +48 giờ nhịn ăn (n = 8 mỗi nghiệm thức nhịn ăn).
3.4. Biến thể giữa và trong phạm vi cá thể
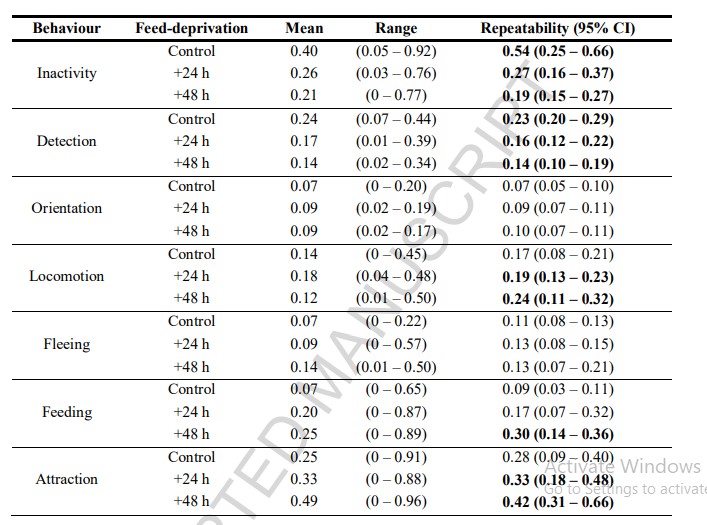
Phân tích khả năng lặp lại đối với các nhóm nhịn ăn cho thấy sự khác biệt đáng kể về tính nhất quán hành vi giữa các nghiệm thức (Bảng 3). Khả năng lặp lại đáng kể đã được tìm thấy trong hai hành vi thụ động được xem xét ở đây, không hoạt động và phát hiện, trên tất cả các nghiệm thức nhịn ăn, với nhóm đối chứng thể hiện khả năng lặp lại cao nhất trong cả hai hành vi. Điều này trái ngược với các hành vi chủ động định hướng và chạy trốn mà không được phát hiện là có thể lặp lại đáng kể trên bất kỳ nhóm nhịn ăn nào. Liên quan đến việc kiếm ăn, chỉ những cá thể trong nhóm nhịn ăn +48 giờ cho thấy khả năng lặp lại đáng kể thời gian dành cho việc tham gia vào hành vi này. Ngoài ra, sự nhất quán về hành vi trong việc hấp dẫn thức ăn và vận động đã được nhìn thấy ở tôm bị bỏ đói trong +24 giờ và +48 giờ (Bảng 3). Những kết quả này về ảnh hưởng của thời gian nhịn ăn được minh họa thêm trong phân tích CV (Hình 4), hỗ trợ các kết quả được trình bày bởi phân tích độ lặp lại. Đối với các hành vi thụ động được kiểm tra, dường như có sự gia tăng sự thay đổi giữa các cá thể ở các giai đoạn nhịn ăn lớn hơn được minh họa bằng cách tăng giá trị CV. Ngược lại, với thời gian nhịn ăn ngày càng tăng, các hành vi tích cực cho thấy sự sụt giảm chung ở giữa và trong các biến thể riêng lẻ, được minh họa bằng dữ liệu CV cho cả việc tìm thức ăn và hấp dẫn.
Bảng 3. Ước tính độ lặp lại cho các hành vi được đo lường theo các nhóm nhịn ăn. Đơn vị tính trung bình là tỷ lệ thời gian dành cho việc thực hiện từng hành vi (n= 8 con tôm mỗi nghiệm thức). Độ lặp lại được tính toán từ các mô hình hiệu ứng hỗn hợp đơn biến với nhận dạng tôm được bao gồm như một hiệu ứng ngẫu nhiên. Khởi động tham số được sử dụng để tính toán khoảng tin cậy (95% CI). Ước tính độ lặp lại đáng kể được in đậm.
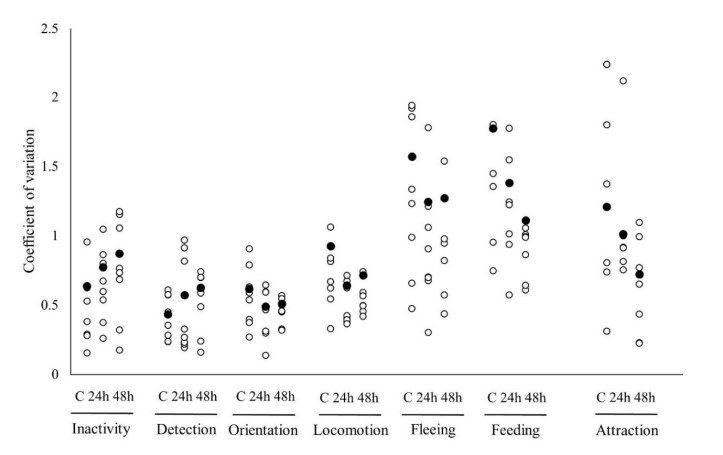
Hình 4. Hệ số thay đổi giữa các cá thể (vòng tròn đen) và trong các cá thể (vòng tròn trắng) đối với thời gian thực hiện các hành vi của trạng thái (không hoạt động, phát hiện, định hướng, vận động, chạy trốn và kiếm ăn) và điểm thu hút thức ăn (%). Dữ liệu dành cho tôm đối chứng (C), +24 giờ và +48 giờ (n= 8 mỗi nghiệm thức, N.B. một số điểm dữ liệu trùng lặp).
Như đã đề cập ở trên, không thể đưa tình trạng lột xác vào phân tích độ lặp lại đầy đủ vì không thể xác định được sự thay đổi trong phạm vi cá thể do thời gian ngắn mỗi con tôm riêng lẻ trong giai đoạn sau lột xác. Tuy nhiên, phân tích sự biến đổi giữa các cá thể (Hình 5) cho thấy sự giảm sự thay đổi trong tất cả các hành vi hoạt động khi tôm ở trạng thái lột xác giữa các cá thể. Mặc dù không có mô hình rõ ràng về ảnh hưởng của tình trạng lột xác đối với CV đối với các hành vi thụ động, tôm trong quá trình lột xác giữa các lần lột xác đã thể hiện giá trị CV cao nhất.
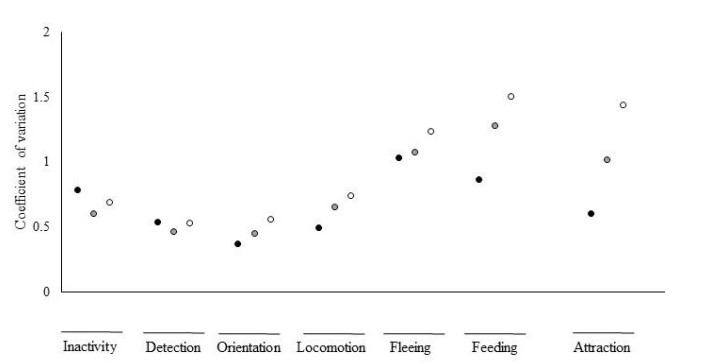
Hình 5. Hệ số thay đổi giữa các cá thể trong quá trình lột xác giữa các cá thể (vòng tròn đen), tiền lột xác (vòng tròn màu xám) và trạng thái sau lột xác (vòng tròn trắng) đối với thời gian thực hiện các hành vi (không hoạt động, phát hiện, định hướng, vận động, chạy trốn và cho ăn) và điểm thu hút thức ăn (n = 24 con tôm mỗi trạng thái lột xác vì mỗi con tôm trải qua cả ba giai đoạn lột xác, do đó các thanh đại diện cho các phép đo lặp đi lặp lại trên cùng một cá thể).
4. Thảo luận
Mặc dù tầm quan trọng của hành vi của động vật trong nuôi trồng thủy sản ngày càng được công nhận, nhưng điều này chưa thực sự được khám phá liên quan đến nuôi tôm. Ở đây kiến thức này đã được bổ sung bằng cách nghiên cứu hành vi kiếm ăn ở tôm thẻ chân trắng L. vannamei. Kết quả chỉ ra rằng tôm thẻ chân trắng L. vannamei điều chỉnh các hành vi và độ hấp dẫn thức ăn để đáp ứng với các giai đoạn nhịn ăn và các giai đoạn khác nhau trong chu kỳ lột xác. Điều này có ý nghĩa đối với sự hiểu biết về hành vi của tôm trong môi trường nuôi trồng thủy sản, nơi các cá thể có khả năng thể hiện sự khác biệt về trạng thái sinh lý.
Trong số ít các nghiên cứu được thực hiện trên tôm thẻ chân trắng L. vannamei, những thay đổi trong hành vi kiếm ăn đã được ghi nhận theo các điều kiện khác nhau. Ví dụ, tôm thẻ chân trắng L. vannamei ăn nhiều hơn trong thời kỳ tối và không hoạt động nhiều hơn trong ánh sáng (Pontes và cộng sự, 2006). Tôm được kiếm ăn 7 lần mỗi ngày (tức là không nhịn ăn), cũng không hoạt động nhiều hơn (Lima và cộng sự, 2009). Trong nghiên cứu này, khi thức ăn có sẵn trong suốt các thử nghiệm hành vi, các hành vi thụ động, năng lượng thấp (ví dụ như không hoạt động, phát hiện) (Da Costa và cộng sự, 2016), đã chiếm ưu thế ở tôm từ nghiệm thức đối chứng và ở trạng thái sau lột xác, nơi quan sát thấy mức độ hoạt động kiếm ăn thấp nhất. Ngoài ra, việc không hoạt động có mối tương quan tiêu cực với việc kiếm ăn trên tất cả các nhóm nhịn ăn và lột xác, với mối tương quan tiêu cực mạnh mẽ hơn ở tôm nhịn ăn hoặc lột xác hơn. Các hành vi thụ động dường như là dấu hiệu đầu tiên hữu ích cho thấy động lực kiếm ăn thấp; nếu tôm có động lực kiếm ăn thì không hoạt động có xu hướng là hành vi đầu tiên giảm khi thức ăn được cung cấp (Silva và cộng sự, 2012). Hơn nữa, tôm có thêm thời gian nhịn ăn (tức là +24 giờ, +48 giờ) và ở trạng thái liên hoặc trước khi lột xác được kiếm ăn nhiều hơn so với tôm đối chứng và sau lột xác. Các thử nghiệm dinh dưỡng trước đây đã cho tôm nhịn ăn trong 18-24 giờ (ví dụ như Holland và Borski, 1993; Sanchez và cộng sự, 2005; Nunes và cộng sự, 2006; Derby và cộng sự, 2016) và / hoặc được sử dụng trong cùng một trạng thái lột xác (ví dụ: Sanchez và cộng sự, 2005; Smith và cộng sự, 2005). Tuy nhiên, điều này không phải lúc nào cũng phổ biến. Các kết quả được trình bày ở đây cho thấy tình trạng nhịn ăn và lột xác là những yếu tố quan trọng cần được kiểm soát đúng cách khi thực hiện các nghiên cứu dinh dưỡng.
Các hành vi sự kiện như được nghiên cứu ở đây (tức là nhịp mắt, nhịp chân, lau râu) chưa được xem xét trong các nghiên cứu trước đây về hành vi của tôm thẻ chân trắng L. vannamei. Kết quả chỉ ra rằng thời gian nhịn ăn không ảnh hưởng đến sự xuất hiện của những hành vi này, tuy nhiên sự khác biệt đáng kể đã được tìm thấy với tình trạng lột xác. Tôm sau khi lột xác cho thấy tần suất nhịp mắt và lau râu cao hơn so với tôm trước và giữa các lần lột xác. Tôm gần đến giai đoạn lột xác (tức là vào cuối giai đoạn trước khi lột xác và sau lột xác) dễ bị căng thẳng hơn (Wajsbrot và cộng sự, 1990) và do đó, có thể sự gia tăng các hành vi này ở tôm sau lột xác là sự phản ánh của căng thẳng. Các quan sát về việc đánh nhịp miệng (ví dụ như nhịp chân như được nghiên cứu ở đây) trước đây đã được ghi nhận ở các loài giáp xác khác như tôm hùm gai Caribbean (Panulirus argus) và có thể được liên kết với hành vi chải chuốt trước đó (AGB) hoặc tự chải chuốt (Barbato và Daniel, 1997; Wroblewska và cộng sự, 2002). Đây là nơi các loài giáp xác làm sạch phần miệng bằng cách sử dụng chân ngực hoặc nhịp chúng một cách mạnh mẽ để loại bỏ các hạt khỏi râu. Mức AGB có thể khác nhau tùy thuộc vào chất hấp dẫn hóa học được sử dụng (ví dụ: Zimmer-Faust và cộng sự, 1984; Barbato và Daniel, 1997; Daniel và cộng sự, 2001). Trong nghiên cứu này, không có sự khác biệt đáng kể nào về tần số của nhịp chân càng được tìm thấy liên quan đến tình trạng nhịn ăn hoặc lột xác, có thể là do chỉ có một loại thức ăn được sử dụng trong quá trình thử nghiệm. Các nghiên cứu trong tương lai có thể tập trung nhiều hơn vào hành vi này để nghiên cứu xem có mối liên hệ tương tự giữa AGB và chất hấp dẫn ở các loài He hay không.
Độ hấp dẫn của thức ăn đã được nghiên cứu trước đây để đánh giá sự lựa chọn tôm giữa các loại thức ăn thử nghiệm khác nhau. Hartati và Briggs (1993) đã đánh giá mức độ hấp dẫn theo số lượng tôm thẻ chân trắng trên khay kiếm ăn có khẩu phần ăn khác nhau và thời gian kiếm ăn. Smith và cộng sự (2005) đã tính toán sự hấp dẫn ở tôm sú châu Á (Penaeus monodon) giữa khẩu phần ăn thử nghiệm khác nhau và khẩu phần ăn cơ bản. Nunes và cộng sự (2006) đã sử dụng một cách tiếp cận tương tự như nghiên cứu này, tính toán định hướng và thời gian vận động mà từng con tôm cần để đến một khu vực chứa thức ăn trong Y-maze cho thấy tôm có sự lựa chọn giữa 2 khẩu phần ăn khác nhau. Trong nghiên cứu này, tình trạng nhịn ăn và lột xác đã ảnh hưởng đáng kể đến độ hấp dẫn thức ăn, với tôm bị nhịn ăn trong +48 giờ và ở trạng thái lột xác cho thấy cách tiếp cận nhanh nhất đối với khu vực kiếm ăn. Tuy nhiên, cách tiếp cận nhanh với khay ăn và tương tác với thức ăn không nhất thiết dẫn đến thời gian kiếm ăn kéo dài, vì tôm có thể ngừng ăn sau khi đến khay. Việc tôm có còn ăn ở khay ăn hay không sẽ phụ thuộc vào hành vi ăn hoặc không ăn như đã được ghi nhận trước đây ở các loài giáp xác khác như tôm càng xanh (Macrobrachium rosenbergii) (Steiner và Harpaz, 1987) và ốc mượn hồn (Pagurus granosimanus) (Wight và cộng sự, 1990). Trong nghiên cứu này, một mối tương quan tích cực đã được tìm thấy giữa hoạt động kiếm ăn và độ hấp dẫn thức ăn, cho thấy tôm đến khay ăn nhanh chóng có động lực hơn để kiếm ăn. Vì chỉ có một loại thức ăn được sử dụng trong nghiên cứu này, nên không biết liệu mối quan hệ giữa hấp dẫn thức ăn và hoạt động kiếm ăn ở tôm thẻ chân trắng L.vannamei có bị ảnh hưởng bởi các loại thức ăn khác hay không. Thật vậy, có thể sử dụng sức mạnh của mối quan hệ này để đánh giá chất lượng thức ăn, nhưng ý tưởng này đòi hỏi phải nghiên cứu thêm. Trong nghiên cứu này, sục khí và lưu lượng nước không có mặt trong lĩnh vực thử nghiệm, phù hợp với các thử nghiệm dinh dưỡng trước đây trên tôm (ví dụ: Pittet và cộng sự, 1996; Sanchez và cộng sự, 2005; Nunes và cộng sự, 2006) vì dòng nước có thể gây ra sự phân tán hóa học cao xung quanh các bể thí nghiệm quy mô nhỏ và tôm phản ứng chủ yếu với dòng chảy, đưa ra hình ảnh không chính xác về khả năng hấp dẫn thức ăn. Do đó, trong nghiên cứu này, có thể các tín hiệu thức ăn đến từng con tôm tùy thuộc vào khả năng tạo ra dòng nước, điều này cũng có thể góp phần vào sự thay đổi cá thể trong phản ứng với thức ăn.
Biến thể của tôm là một cân nhắc quan trọng cho các nghiên cứu hành vi (Briffa và Sneddon, 2016; Hewes và Chaves-Campos, 2018) vì nó có thể ảnh hưởng đến khả năng tạo ra kết quả không thuyết phục trong một nghiên cứu (Briffa và cộng sự, 2008; Dingemanse và Dochtermann, 2013). Trong nghiên cứu này, sự thay đổi của tôm liên quan đến nhịn ăn đã được nghiên cứu bằng cách sử dụng phân tích độ lặp lại. Các hành vi thụ động trở nên kém nhất quán hơn (tức là giá trị lặp lại thấp hơn) và các hành vi chủ động (ví dụ: kiếm ăn) trở nên nhất quán hơn (tức là giá trị lặp lại cao hơn) vì tôm bị nhịn ăn lâu hơn. Những phát hiện này được hỗ trợ thêm bởi các kết quả thu được từ phân tích CV cho thấy xu hướng giảm sự thay đổi giữa (đối với cả tình trạng nhịn ăn và lột xác) và trong tôm (chỉ dành cho nhịn ăn) trong các hành vi hoạt động khi tôm ở trạng thái lột xác và nhịn ăn nhiều hơn. Nghiên cứu trước đây về tính nhất quán hành vi đã tìm thấy những phát hiện tương tự ở các loài giáp xác khác. Ví dụ, ốc mượn hồn (Pagurus bernhardus) (Briffa và cộng sự, 2008) và cua càng to (Heterozius rotundifrons) (Hazlett và Bach, 2009) đã được thử nghiệm nhiều lần để xác định biến thể cá nhân liên quan đến hành vi phòng thủ của động vật ăn thịt. Các cá thể khác nhau trong hành vi phòng thủ nhưng thể hiện mức độ nhất quán cá thể cao trong các điều kiện. Alexander và cộng sự (2015) đã tìm thấy sự giảm sự thay đổi giữa các cá thể trong hành vi kiếm ăn ở động vật lưỡng cư biển (Echinogammarus marinus) khi các cá thể bị nhịn ăn lâu hơn. Tuy nhiên, sự thay đổi trong phạm vi cá thể tăng lên với thời gian nhịn ăn lâu hơn. Trong L.vannamei, Santos và cộng sự (2016) nhận thấy rằng sự nhất quán hành vi trong vận động và hoạt động kiếm ăn được thúc đẩy bởi nhịp sinh học. Hơn nữa, nghiên cứu này phản ánh tình trạng nhịn ăn và lột xác ảnh hưởng đến sự biến đổi cá thể như thế nào, với những tác động quan trọng đối với các nghiên cứu hành vi trong tương lai. Sự hiện diện của khả năng lặp lại trong một số hành vi được đo lường trong nghiên cứu này cho thấy sự tồn tại của tính cách động vật trong L. vannamei (Briffa và Weiss, 2010), một thuật ngữ giải thích những khác biệt nhất quán này trong hành vi cá thể. Khái niệm này đã được đề cập trong các loài giáp xác khác (Briffa và cộng sự, 2008; Bridger và cộng sự, 2015; Hewes và Chaves-Campos, 2018) nhưng không có trong tôm He, và do đó mở ra con đường thú vị cho nghiên cứu trong tương lai.
5. Kết luận
Các kết quả được trình bày ở đây cho thấy tình trạng nhịn ăn và lột xác có ảnh hưởng đáng kể đến hành vi kiếm ăn của tôm thẻ chân trắng với những tác động quan trọng đối với việc thiết kế các thử nghiệm lựa chọn thức ăn và hành vi trong tương lai. Mức độ kiếm ăn cao nhất và giảm mức độ biến đổi cá thể đã được tìm thấy ở tôm bị nhịn ăn thêm 48 giờ và trong quá trình lột xác. Các hành vi thụ động (tức là không hoạt động, phát hiện) tương quan với động lực kiếm ăn thấp. Sử dụng khẩu phần ăn tốt, điểm thu hút thức ăn là một phương pháp hiệu quả để đánh giá mức độ hoạt động kiếm ăn và cũng bị ảnh hưởng bởi tình trạng nhịn ăn và lột xác. Ngoài ra, khả năng lặp lại của một số hành vi được phân tích có thể cung cấp bằng chứng về tính cách động vật ở tôm He, mặc dù cần phải nghiên cứu thêm. Tầm quan trọng của việc xem xét tình trạng nhịn ăn và lột xác đã được xác định khi nghiên cứu hành vi kiếm ăn của tôm thẻ chân trắng, cung cấp thông tin cơ bản quan trọng trong việc phát triển việc sử dụng hành vi để cải thiện sản lượng tôm thẻ chân trắng.
Theo Guillermo Bardera, Matthew A.G. Owen, Daniel Pountney, Mhairi E. Alexander, Katherine A. Sloman
Nguồn: https://sci-hub.hkvisa.net/10.1016/j.aquaculture.2019.734222
Biên dịch: Nguyễn Thị Quyên – Bình Minh Capital
Xem thêm:
- Phần 1: Giun Nhiều Tơ (Perinereis helleri) Nuôi Trong Hệ Thống Lọc Cát Giúp Tận Dụng Chất Dinh Dưỡng Từ Nước Thải Ao Nuôi Tôm Sú (Penaeus monodon), Nhưng Mang Rủi Ro Về Mầm Bệnh IHHNV
- Phần 2: Giun Nhiều Tơ (Perinereis helleri) Nuôi Trong Hệ Thống Lọc Cát Giúp Tận Dụng Chất Dinh Dưỡng Từ Nước Thải Ao Nuôi Tôm Sú (Penaeus monodon), Nhưng Mang Rủi Ro Về Mầm Bệnh IHHNV
- Tôm Sạch Mầm Bệnh: Tiến Bộ Của Ngành Nuôi Tôm