
 English
English
Kỹ Thuật Nuôi, Tin tức
Đánh Giá Đặc Điểm Độc Lực Và Kháng Kháng Sinh Của Vibrio Phân Lập Từ Tôm Thẻ Chân Trắng (Penaeus vannamei) Trong Hệ Thống Nuôi Ở Miền Nam Trung Quốc
Điểm nổi bật
- Sự phân bố quần thể, độc lực và nguy cơ kháng kháng sinh của Vibrio trong hệ thống nuôi tôm ở Trung Quốc lần đầu tiên được đánh giá.
- Vibrio phân lập từ hệ thống nuôi tôm là đại diện cho sự đa dạng di truyền và trao đổi chất điển hình.
- Đánh giá rủi ro của Vibrio trong hệ thống thủy sinh cho thấy rủi ro cuối cùng đối với hệ sinh thái thủy sinh và các khía cạnh sức khỏe cộng đồng.
Tóm tắt
Bệnh do vi khuẩn Vibrio gây ra trong quá trình ương nuôi tôm có nguy cơ lây lan sang các vùng nuôi thủy sản khác nhau thông qua việc vận chuyển ấu trùng giữa các vùng khác nhau. Do đó, sự phân bố quần thể và nguy cơ độc lực, kháng kháng sinh của 5 loài Vibrio gây bệnh trong hệ thống nhân giống tôm (Penaeus vannamei) ở Trung Quốc lần đầu tiên được đánh giá. Tổng cộng có 418 phân lập được thu hồi từ mẫu tôm, nước nuôi và mồi sinh học, và 312 phân lập được xác định là các loài trong hệ thống nuôi tôm ở Trung Quốc cho thấy không có hoặc có áp lực chọn lọc thấp đối với khả năng kháng cefoperazone/sulbactam, enrofloxacin, ciprofloxacin, fluoroquine, florfenicol, tetracycline và doxycycline. Nhưng chỉ có 5 gen kháng thuốc được phát hiện với (8,8%) và kiểu gen kháng kháng sinh trước đây không tương quan với kiểu hình của chúng. Các kết quả nghiên cứu có liên quan cung cấp cơ sở lý thuyết cho việc theo dõi dịch bệnh trong hệ thống thủy sinh ở Trung Quốc và nhắm tới rủi ro cuối cùng đối với hệ sinh thái thủy sinh và các khía cạnh sức khỏe cộng đồng.
1. Giới thiệu
Nuôi tôm là một trong những ngành nuôi trồng thủy sản quan trọng đóng góp cho nền kinh tế toàn cầu và phụ thuộc nhiều vào môi trường chăn nuôi, tôm thẻ chân trắng được coi là loài nuôi quan trọng nhất trên thế giới với sản lượng hàng năm đạt 5 triệu tấn (FAO, 2016). Dịch bệnh bùng phát do vi khuẩn, đặc biệt là bệnh do vi khuẩn Vibrio gây ra đang trở thành yếu tố quan trọng hạn chế sản lượng nuôi tôm ổn định. Và Vibrio có thể gây tỷ lệ chết lên đến 100% ở động vật thủy sinh (Kumar và cộng sự, 2014). Là yếu tố quan trọng nhất để sản xuất tôm nuôi ổn định, ngành nuôi tôm phụ thuộc rất nhiều vào việc quản lý sức khỏe của chương trình chăn nuôi thực hành nuôi ấu trùng. Các loài Vibrio spp., chẳng hạn như V. harveyi, V. parahaemolyticus, V. alginolyticus và V. cholerae đã được báo cáo là gây bệnh trong quá trình nhân giống tôm (Haldar và cộng sự, 2007, Zou và cộng sự, 2020). Những vi khuẩn này làm tăng mạnh nguy cơ nuôi tôm và truyền mầm bệnh vào môi trường.
Vibriosis được tạo điều kiện thuận lợi bởi nhiều yếu tố độc lực, cho phép mầm bệnh lây nhiễm và gây hại cho vật chủ và cuối cùng thoát ra khỏi vật chủ bị nhiễm bệnh (Schroeder và cộng sự, 2017). Đặc điểm độc lực bao gồm sự tiết chitinase, giống myovirus, giống siphovirus và haemolysin được biểu hiện ở V. harveyi (Ruwandeepika và cộng sự, 2010). Biểu hiện của độc tố cholix (chx), độc tố tả (ctx), độc tố ruột ổn định nhiệt (stn) và neuraminidase ở V. cholerae (Awasthi và cộng sự, 2019, Gennari và cộng sự, 2012). Các gen liên quan đến hemolysin (tlh, tdh và trh) ở V. parahaemolyticus nằm trong số các gen độc lực của Vibrio (Bej và cộng sự, 1999). Hơn nữa, một số gen độc lực (PirAvp /PirBvp) nằm trong plasmid có thể lây lan giữa các loài Vibrio khác nhau (Sirikharin và cộng sự, 2015). Sự giao tiếp giữa các phân tử tín hiệu cũng góp phần vào việc giao tiếp độc lực dưới vi khuẩn gây bệnh (De Kievit và Iglewski, 2000). Việc phân tích các gen độc lực chính của vi khuẩn Vibrio phân lập từ môi trường nước có thể cung cấp những hiểu biết có ý nghĩa về cơ chế bệnh sinh của bệnh Vibrio và giúp thiết lập các biện pháp phòng ngừa hiệu quả và chuyên sâu.
Nói chung, các loài Vibrio được công nhận là rất nhạy cảm với hầu hết các loại thuốc kháng sinh (Daniels và Shafaie, 2000, Elmahdi và cộng sự, 2016). Trong một số trường hợp nhất định, sự phát triển của nhiều chủng phân lập kháng thuốc và sự hiện diện rộng rãi của các yếu tố kháng thuốc trong quần thể vi khuẩn có thể phản ánh đặc điểm kiểu hình kháng thuốc của vi khuẩn trong những trường hợp nhất định (Reboucas và cộng sự, 2011). Hiện tượng kháng kháng sinh được coi là mối đe dọa toàn cầu quan trọng đối với hệ sinh thái môi trường, sức khỏe cộng đồng và an toàn thực phẩm (FAO, 2016). Hơn nữa, việc sử dụng kháng sinh trong hệ thống nuôi trồng thủy sản có thể là nguyên nhân dẫn đến sự phân bố rộng rãi các gen kháng kháng sinh (Igbinosa, 2016). Các gen mã hóa tính kháng β-lactam như penA, blaTEM-1 (Letchumanan và cộng sự, 2015), ampC (Jacoby, 2009), các gen kháng chloramphenicol như floR, catI, và catII và một số gen tets (Letchumanan và cộng sự, 2015), đã được xác định ở Vibrio. Các gen kháng di động như qnrVC và qnrS cũng đã được hình thành (Fonseca và cộng sự, 2008). Thách thức lớn nhất trong ngành nuôi trồng thủy sản là phương pháp phù hợp để phòng ngừa và kiểm soát dịch bệnh thủy sản. Phân tích khả năng kháng kháng sinh của mầm bệnh ở một khu vực cụ thể là một lĩnh vực quan trọng cho các chương trình kiểm soát dịch bệnh hiệu quả và phát triển thân thiện với môi trường.
Việc chuyển con giống sang các khu vực và giai đoạn khác nhau càng làm tăng thêm nguy cơ lây truyền mầm bệnh. Sự phát triển bền vững của ngành nuôi tôm phụ thuộc rất nhiều vào việc quản lý sức khỏe của các hoạt động nuôi ấu trùng. Dựa trên những thực tế này, nghiên cứu này được thực hiện để phân tích sự phân bố quần thể, đặc điểm độc lực và nguy cơ kháng khuẩn của Vibrio spp. ở các giai đoạn nuôi tôm khác nhau. Các kết quả này có thể dùng làm dữ liệu cơ bản để theo dõi dịch bệnh trong các hệ thống nhân giống thủy sản và xác định rủi ro cuối cùng đối với hệ sinh thái thủy sinh và các khía cạnh sức khỏe cộng đồng.
2. Vật liệu và phương pháp
2.1. Địa điểm thử nghiệm
Các khu vực ven biển phía nam, bao gồm tỉnh Hải Nam và Quảng Đông, là những khu vực nuôi tôm chính ở Trung Quốc, với nhiệt độ trung bình hàng năm dao động từ 22°C đến 26°C (Xu và cộng sự, 2017). Những địa điểm đó thích hợp cho việc ương nuôi tôm giống lâu năm và quy trình nuôi tôm công nghiệp quy mô lớn chính ở Trung Quốc tập trung ở các tỉnh này, sau đó con giống được vận chuyển khắp đất nước từ khu vực này (Yang và cộng sự, 2022).
2.2. Thu thập mẫu
Có 5 con tôm bố mẹ được chọn ngẫu nhiên từ bể ương, 1 g trứng tôm được thu từ bể ấp và 1-2 g ấu trùng được thu ngẫu nhiên từ bể ương của ấu trùng ở các giai đoạn khác nhau, bao gồm nauplius, zoea, mysis và tôm post bằng lưới lụa đã khử trùng và 500 mL mẫu nước sinh sản có liên quan được thu thập đồng thời. Mồi sinh học (giun, mực đông lạnh, vi tảo và artemia) cho các giai đoạn phát triển cũng được thu thập ở mức 1–2 g.
Trứng tôm, ấu trùng ở các giai đoạn khác nhau và mẫu mồi sinh học được rửa nhiều lần bằng dung dịch NaCl 1,5% vô trùng trong 3-5 lần để loại bỏ vi sinh vật bám trên bề mặt. Sau đó, mẫu được làm khô bằng gạc vô trùng và đưa về phòng thí nghiệm ở nhiệt độ thấp để phân tích tiếp. Tôm bố mẹ được mổ xẻ trong điều kiện vô trùng, gan tụy và ruột được lấy mẫu.
2.3. Nuôi cấy vi khuẩn và phân lập Vibrio
Các mẫu tôm nặng 0,1 g (tôm bố mẹ, trứng tôm, ấu trùng, zoea, mysis và tôm post) và mồi sinh học (giun, mực đông lạnh, vi tảo và artemia) được cân từng loại để nghiền vô trùng và pha loãng gradient 10 lần bằng vô trùng. Dung dịch NaCl 1,5%. Các mẫu nước sinh sản từ bể nuôi tương ứng được lấy để pha loãng gradient 10 lần thông qua dung dịch NaCl 1,5% vô trùng. Sau đó, 100 µL nước pha loãng của mỗi mẫu được phết đều trên các đĩa thạch thiosulphate citrate muối mật sucrose (TCBS, LAND BRIDGE, Trung Quốc) để nuôi cấy Vibrio, ủ ở 28°C trong 24 giờ. Theo hình thái khuẩn lạc, màu sắc khuẩn lạc, kích thước khuẩn lạc và các đặc điểm khác, các khuẩn lạc khác nhau được chọn và phết trên các đĩa thạch TCBS để làm sạch vi khuẩn.
2.4. Xác định các chủng Vibrio phân lập
Các chủng vi khuẩn đã được tinh chế được cấy vào đĩa thạch trypticase đậu nành (TSB, MDBio, Inc, Trung Quốc) được bổ sung 1,5% NaCl và nuôi cấy ở 28°C trong 24 giờ. Trong điều kiện vô trùng, các khuẩn lạc đơn lẻ được chọn và treo lại trong 100 µL nước khử ion vô trùng. Sau 15 phút ngâm trong bể kim loại ở 99°C và ly tâm ở 5000 g trong 5 phút ở 4°C, phần nổi phía trên được sử dụng làm mẫu DNA. Nồng độ DNA ở phần nổi phía trên được đo bằng máy quang phổ (NanoDrop1000, Thermo Scientific, USA) và điều chỉnh ở khoảng 100 ng/µL bằng nước khử ion vô trùng, được sử dụng làm DNA mẫu PCR.
Đầu tiên, việc xác nhận danh tính chi của các chủng Vibrio giả định được phân lập dựa trên các đoạn mồi phổ quát dành riêng cho việc xác định rDNA ở thập niên 16 (Byers và cộng sự, 1998). Sau đó, các mồi dành riêng cho loài được sử dụng để phân loại các chủng phân lập thành V. alginolyticus, V. harveyi, V. parahaemolyticus, V. cholerae và V. campbellii với các mồi cụ thể như đã mô tả trước đây (Zhou và cộng sự, 2007, Pang và cộng sự, 2006, Tarr và cộng sự, 2007, Nandi và cộng sự, 2000, Haldar và cộng sự, 2010). Dữ liệu trình tự được so sánh với cơ sở dữ liệu NCBI (https://www.ncbi.nlm.nih.gov/) bằng BLAST. Các gen mục tiêu, trình tự mồi, kích thước dải và nhiệt độ ủ được trình bày trong Bảng bổ sung 1.
2.5. Đặc tính phân tử của các gen độc lực dưới các chủng Vibrio phân lập
Tổng cộng có 20 gen độc lực đã được phát hiện, bao gồm tlh, tdh, trh, rpoS và VAC ở V. alginolyticus (Bej và cộng sự, 1999, Huang và cộng sự, 2019, Di Pinto và cộng sự, 2005); tlh, tdh, trh, VpadF và pirA/B ở V. parahaemolyticus (Bej và cộng sự, 1999, Liu và Chen, 2015, Dangtip và cộng sự, 2015); luxR, chiA, vhml, vhs và vhh ở V. harveyi (Ruwandeepika và cộng sự, 2010); ctxA, stn, chxA, cqsA và nanH ở V. cholerae (Fields và cộng sự, 1992, Awasthi và cộng sự, 2019, Kumar và cộng sự, 2021, Gennari và cộng sự, 2012); luxR, chiA, sepro, flaA và vch ở V. campbellii (Ruwandeepika và cộng sự, 2010, Yang và Defoirdt, 2015, Haldar và cộng sự, 2010). Mỗi phản ứng bao gồm biến tính sơ bộ ở 94°C trong 5 phút, kèm theo 30 chu kỳ, mỗi chu kỳ bao gồm biến tính ban đầu ở 94°C trong 60 giây, sau đó ủ ở nhiệt độ thay đổi trong 60 giây và các bước mở rộng trong 60 giây trong khi phản ứng cuối cùng độ giãn dài là 72°C trong 5 phút. Các mồi cho các gen độc lực và nhiệt độ ủ của chúng được trình bày trong Bảng bổ sung 2.
2.6. Đặc tính phân tử của các gen kháng kháng sinh dưới các chủng Vibrio phân lập
Phân lập Vibrio với các mồi đặc hiệu được thực hiện để phát hiện sự hiện diện hay vắng mặt của 23 gen kháng kháng sinh thuộc 8 loại. Các gen kháng kháng sinh mục tiêu là: gen kháng β-lactam (blaCMY, blaPSE và ampC) (Moffat và cộng sự, 2020, Qiao và cộng sự, 2017, Gxalo và cộng sự, 2021), gen kháng aminoglycoside (strA và strB) (Chuanchuen và Padungtod, 2009), gen kháng macrolide (MphA, ermA và ermC) (Nguyen và cộng sự, 2009, Spiliopoulou và cộng sự, 2004), gen kháng quinolones (QnrVC, oqxB và soxR) (Liu và cộng sự, 2013, Kim và cộng sự, 2009, Komp Lindgren và cộng sự, 2003), gen kháng sulfonamid (sul1, sul2 và sulA) (Yuan và cộng sự, 2021, Pei và cộng sự, 2006), gen kháng chloramphenicol (cat-2 và cfr) (Jiang và cộng sự, 2013, Kehrenberg và Schwarz, 2006), gen kháng Tetracyclines (tetC, tetE và tetM) (Jun và cộng sự, 2004, Kim và cộng sự, 2004), và integron (Int1, Int2, Int3 và Int4) (Koeleman và cộng sự, 2001, Xu và cộng sự, 2007, Rodriguez-Blanco và cộng sự, 2012). Trình tự mồi, kích thước khuếch đại và nhiệt độ ủ của mồi được trình bày trong Bảng bổ sung 3.
2.7. Kiểu hình kháng kháng sinh
Phương pháp khuếch tán đĩa tiêu chuẩn (xét nghiệm Kirby-Bauer) được thực hiện để phân tích độ nhạy cảm với kháng sinh như được mô tả trước đây bởi Bauer và cộng sự (1966) với những thay đổi nhỏ. Tổng cộng có 16 loại kháng sinh được lựa chọn như sau: ampicillin (AMP, 10μg), cefalexin (LEX, 30μg), cefoperazone/sulbactam (CSL, 75/75μg), streptomycin (STR, 10μg), neomycin (NEO, 30μg), erythromycin (ERY, 15μg), clarithromycin (CLR, 15μg), enrofloxacin (ENR, 10μg), ciprofloxacin (CIP, 5μg), flumequine (UB, 30μg), sulfamethoxazole/trimethoprim (SXT, 23,75/1,25µg), sulfadiazine (SDI, 300µg), flufenicol (FFC, 30µg), thiamphenicol (THI, 30µg), tetracycline (TET, 30 µg), doxycycline (DOX, 30µg). Tóm lại, nuôi cấy vi khuẩn được tạo huyền phù trong dung dịch natri clorua 1,5% vô trùng và sau đó phủ lên các đĩa thạch TSB. Đường kính của vùng ức chế được đo sau khi ủ ở 28°C trong 24 giờ và kết quả được giải thích theo hướng dẫn của Viện Tiêu chuẩn Lâm sàng và Phòng thí nghiệm (CLSI, 2018). Các loại kháng sinh và tiêu chí diễn giải đường kính vùng được sử dụng trong nghiên cứu này được tóm tắt trong Bảng bổ sung 4.
2.8. Phân tích chỉ số đặc tính kháng kháng sinh
Phân tích chỉ số kháng kháng sinh (MAR) được sử dụng để xác định mức độ ô nhiễm kháng sinh của vi khuẩn Vibrio (Osundiya và cộng sự, 2013). Các chủng phân lập có chỉ số MAR < 0,2 được cho là có nguồn gốc từ phục hồi từ các nguồn ô nhiễm kháng sinh có nguy cơ thấp trong khi các chủng phân lập có chỉ số MAR > 0,2 là từ các nguồn có nguy cơ cao (Krumperman, 1983). Đồng thời, mức độ phong phú của mô hình kháng kháng sinh (ARPA), với giá trị cao hơn biểu thị mức độ biến đổi cao hơn trong mô hình kháng kháng sinh của các loài Vibrio, đã được tính toán (Deng và cộng sự, 2020). Ngoài ra, chỉ số kháng thuốc (RQ) được sử dụng để xác định những thay đổi có thể có trong kiểu hình kháng kháng sinh của các loài Vibrio khác nhau trong các chương trình nhân giống tôm trên nhiều loại kháng sinh được thử nghiệm (Adesiyan và cộng sự, 2021).
2.9. Phân tích liên kết
Sau đó, mối liên quan giữa kiểu hình kháng kháng sinh và gen kháng kháng sinh đã được tính toán bằng cách sử dụng xét nghiệm Chi bình phương ở cấp chi và các mối liên quan có ý nghĩa thống kê được thể hiện dưới dạng giá trị Cramer để thể hiện cường độ liên kết (Nasaj và cộng sự, 2020). Giá trị của Cramer có phạm vi từ 0 đến 1, trong đó 1 biểu thị sự liên kết hoàn hảo và 0 biểu thị không có mối quan hệ nào. Việc giải thích giá trị của Cramer được thực hiện trong đó các mối liên quan được phân loại là không (0), yếu (>0 đến <0,1), thấp (≥0,1 đến <0,3), trung bình (≥ 0,3 đến <0,5) và mạnh (≥0,5) (Sony và cộng sự, 2021). Kiểm định Chi bình phương được thực hiện bằng phần mềm SPSS phiên bản 16. Giá trị P < 0,05 được coi là có ý nghĩa thống kê.
2.10. Phân tích thống kê
Excel phiên bản 2021 được dùng để tính chỉ số MAR, ARPA và RQ của các loài Vibrio khác nhau. Sử dụng các công cụ có liên quan trong nền tảng đám mây omicshare (https://www.omicshare.com/) để tạo các bản đồ xiếc và nhiệt. Ngoài ra, phiên bản excel 2021 còn được sử dụng để vẽ biểu đồ thanh và biểu đồ hình tròn tổng hợp.
3. Kết quả
3.1. Tỷ lệ và phân bố của các loài Vibrio trong môi trường nuôi tôm
Tổng cộng có 418 chủng được phân lập từ tất cả các mẫu và 312 (74,6%) phân lập được xác nhận thuộc chi Vibrio dựa trên rDNA 16s, trong đó V. alginolyticus, V. harveyi, V. parahaemolyticus, V. cholerae và V. campbellii là loài chiếm ưu thế. Việc xác định thêm dựa trên các mồi đặc trưng cho loài, kết quả như sau: 59 được xác nhận là V. alginolyticus, 35 phân lập V. harveyi, 16 phân lập V. parahaemolyticus, 16 phân lập V. cholerae, 11 phân lập V. campbellii, tổng cộng 137 Vibrio phân lập (Hình bổ sung 1). Sự phân bố của 137 chủng Vibrio phân lập này trong các mẫu tôm, nước giống và mồi sinh học lần lượt là 63, 49 và 25.
V. alginolyticus, V. harveyi, V. parahaemolyticus và V. cholerae được tìm thấy trong tôm, nước ương giống và mồi sinh học, trong khi V. campbellii chỉ được tìm thấy trong tôm và nước ương giống (Hình 1). Tỷ lệ phân bố của V. alginolyticus trong tôm, nước giống và mồi sinh học lần lượt là 28,6% (18/63), 55,1% (27/49) và 56% (14/25), V. harveyi là 22,2% (14/63), 28,6% (14/49) và 28% (25/7), V. parahaemolyticus là 20,6% (13/63), 4,1% (2/49) và 4% (1/25), V. cholerae là 12,7% (8/63), 10,2% (5/49) và 12% (3/25). Ngoài ra, tỷ lệ phân bố của V. campbellii trong tôm và nước nuôi là 15,9% (10/63) và 2,0% (1/49). V. alginolyticus là loài phổ biến nhất với tần suất xuất hiện cao nhất ở tôm, nước giống và mồi.
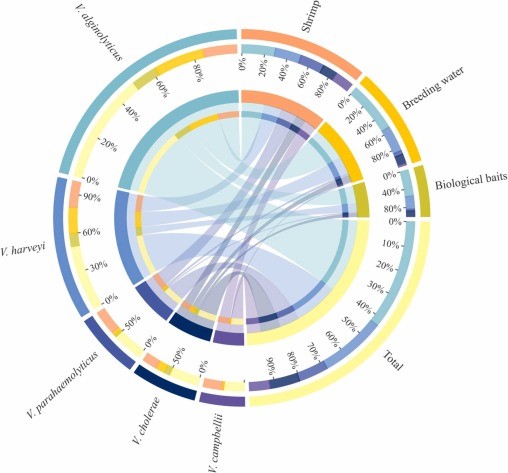
Hình 1. Sự phổ biến của các loài Vibrio trong các chương trình nuôi tôm. Nửa vòng tròn bên trái tượng trưng cho tỷ lệ loài Vibrio có trong mẫu, nửa vòng tròn bên ngoài tượng trưng cho mẫu; màu sắc của vòng tròn bên trong tượng trưng cho các loài Vibrio khác nhau và chiều dài tượng trưng cho tỷ lệ phân bố của các loài Vibrio trong mẫu tương ứng. Nửa vòng tròn bên phải thể hiện tỷ lệ phân bố của các mẫu ở các loài Vibrio khác nhau, vòng tròn bên ngoài thể hiện các loài Vibrio, màu sắc của hình tròn bên trong thể hiện các mẫu khác nhau và chiều dài thể hiện tỷ lệ phân bố của mẫu ở một loài Vibrio nhất định. Giá trị tỷ lệ biểu thị tỷ lệ của nhãn màu tương ứng. Và vạch ở giữa biểu thị sự có mặt của loài Vibrio trong mẫu, đoạn vạch càng dày thì hàm lượng loài Vibrio càng phong phú.
3.2. Đánh giá rủi ro gen độc lực của 5 loại Vibrio
Tổng cộng 10/20 gen độc lực đã được phát hiện trong số 5 loài Vibrio được phân lập từ môi trường sinh sản P. vannamei (Hình bổ sung 2), và sự phân bố các yếu tố quyết định độc lực giữa các loài Vibrio được trình bày trong Hình 2. Đối với V. alginolyticus, phát hiện được 2 trong số 5 gen độc lực, 94,9% V. alginolyticus chứa VAC và phân bố trong các mẫu tôm, nước giống và mồi sinh học lần lượt là 30,5%, 44,1% và 20,3%; 89,8% chủng phân lập có rpoS với sự phân bố trong mẫu tôm, nước giống và mồi sinh học lần lượt là 27,1%, 42,4% và 20,3%. Đối với V. harveyi, 3 trong số 5 gen độc lực đã được phát hiện, tất cả các chủng V. harveyi phân lập đều biểu hiện chiA với sự phân bố trong các mẫu tôm, nước giống và mồi sinh học lần lượt là 40%, 40% và 20%; 11,4% chủng V. harveyi phân lập cho thấy luxR phân bố ở tôm và mức phân bố luxR là 5,7% ở cả mẫu tôm và mẫu nước nuôi; 80% hiển thị vhh với sự phân bố trong các mẫu tôm, nước giống và mồi sinh học lần lượt là 25,7%, 34,3% và 20%. Đối với V. parahaemolyticus, chỉ có tlh dương tính trong số 5 gen độc lực được phát hiện, với tỷ lệ phát hiện là 93,8%. Nó được phân bố trong các mẫu tôm, nước nuôi và mồi sinh học với tỷ lệ lần lượt là 75%, 12,5% và 6,3%. Đối với V. cholerae, phát hiện 1 trong 5 gen độc lực, 68,8% thể hiện chx với sự phân bố trên các mẫu tôm, nước giống và mồi sinh học lần lượt là 37,5%, 18,8% và 12,5%. Phát hiện được tất cả 5 gen độc lực của V. campbellii, 100% V. campbellii phân lập chiA, luxR, sepro và flaA với sự phân bố trong các mẫu tôm và nước nuôi lần lượt là 90,9% và 9,1%; 27,3% chủng phân lập được chứng minh là vch chỉ phân bố trong các mẫu tôm.
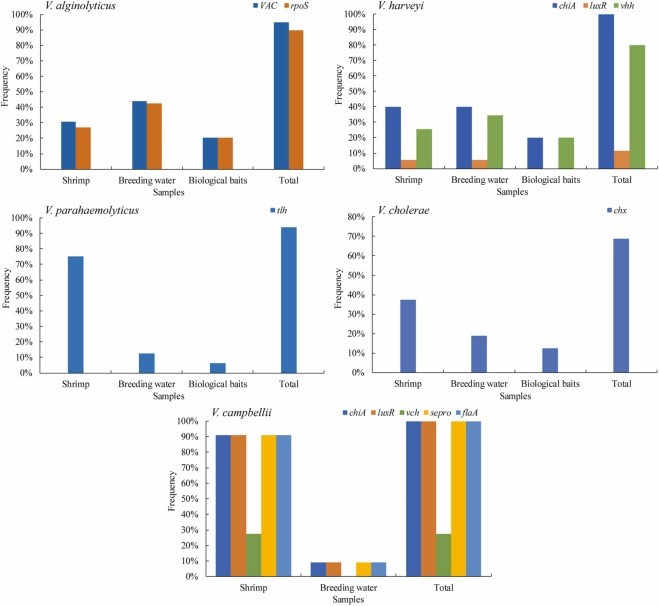
Hình 2. Tỷ lệ gen độc lực của 5 loài Vibrio từ các mẫu khác nhau trong chương trình nhân giống tôm ở Trung Quốc.
3.3. Đặc điểm kiểu hình kháng kháng sinh của 5 loài Vibrio
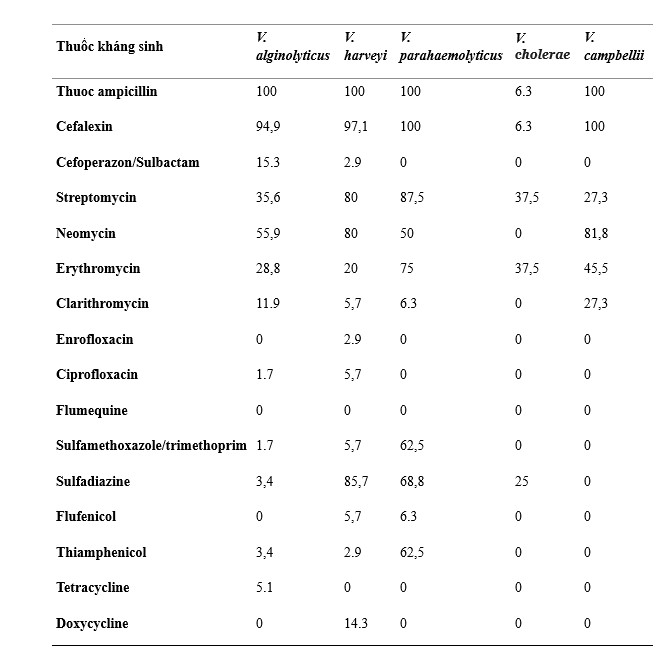
Tất cả các chủng Vibrio phân lập đều có khả năng kháng cao (>50%) với ampicillin (89,1%), cefalexin (86,1%), streptomycin (52,6%) và neomycin (56,9%); tỷ lệ kháng vừa phải (10–50%) với erythromycin (34,3%) và sulfadiazine (34,3%); và tỷ lệ kháng thấp (< 10%) với cefoperazone/sulbactam (7,3%), clarithromycin (9,5%), enrofloxacin (0,7%), ciprofloxacin (2,2%), sulfamethoxazole/trimethoprim (9,5%), flufenicol (2,2%), thiamphenicol (9,5%), tetracycline (2,2%), doxycycline (3,6%) và flumequine (0%) (Hình 3).
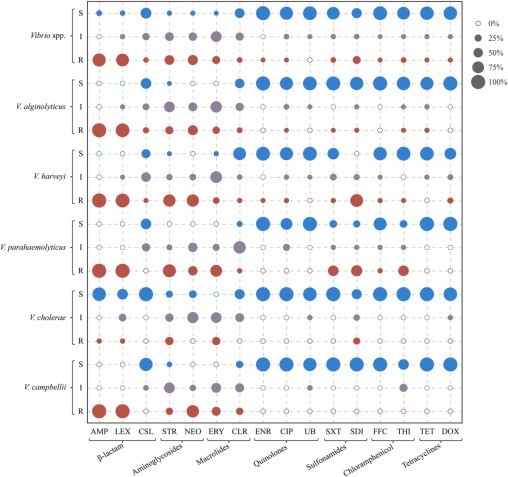
Hình 3. Phân bố độ nhạy cảm với kháng sinh của 5 loài Vibrio. AMP: ampicillin; LEX: cefalexin; CSL: cefoperazon/sulbactam; STR: streptomycin; NEO: neomycin; ERY: erythromycin; CLR: clarithromycin; ENR: enrofloxacin; CIP: ciprofloxacin; UB: flumequine; SXT: sulfamethoxazole/trimethoprim; SDI: sulfadiazine; FFC: flufenicol; THI: thiamphenicol; TET: tetracycline; DOX: doxycycline.
Đối với các loài Vibrio khác nhau, V. alginolyticus, V. harveyi, V. parahaemolyticus và V. cholerae có khả năng kháng ampicillin cao (100%), ceshalexin (94,9–100%) và neomycin (50–81,8%), trong khi V. cholerae cho thấy sức đề kháng thấp (0–6,3%). V. parahaemolyticus cho thấy tính kháng cao với sulfamethoxazole/trimethoprim (62,5%) và thiamphenicol (62,5%), V. alginolyticus, V. harveyi, V. cholerae và V. campbellii có tính kháng thấp (0–5,7%). V. harveyi và V. parahaemolyticus cho thấy khả năng kháng streptomycin cao (80–87,5%), trong khi V. alginolyticus, V. cholerae và V. campbellii cho thấy khả năng kháng vừa phải (27,3–37,5%). V. harveyi, V. parahaemolyticus, V. cholerae và V. campbellii biểu hiện khả năng kháng thấp với cefoperazone/sulbactam (0–2,9%), nhưng V. alginolyticus cho thấy khả năng kháng vừa phải (15,3%). Tương tự, V. alginolyticus, V. parahaemolyticus, V. cholerae và V. campbellii biểu hiện tính kháng thấp với doxycycline (0%), nhưng V. harveyi cho thấy tính kháng trung bình (14,3%). Và cả 5 loài Vibrio đều biểu hiện khả năng kháng enrofloxacin thấp (0–2,9%), ciprofloxacin (0–5,7%), flumequine (0%), flufenicol (0–6,3%), tetracycline (0–5,1%).
3.4. Chỉ số và mô hình của nhiều kiểu hình kháng kháng sinh ở các chủng Vibrio phân lập
Có 38 mẫu kháng kháng sinh trên 137 chủng Vibrio phân lập, trong đó AMP-LEX-SDI, AMP-LEX-STR-NEO-SDI và AMP-LEX là các mẫu chính được thể hiện với tỷ lệ lần lượt là 12,4%, 11,7% và 10,9% trên tổng số chủng phân lập, (Bảng 1). V. alginolyticus có số lượng mẫu kháng kháng sinh cao nhất (26) và mẫu chiếm ưu thế là AMP-LEX và AMP-LEX-SDI. V. harveyi (12), V. parahaemolyticus (9) và V. campbellii (5) đã được theo dõi, các kiểu kháng kháng sinh chính của chúng là AMP-LEX-STR-NEO-SDI, AMP-LEX-STR-NEO-SDI và AMP -LEX-SDI, tương ứng. V. cholerae có ít kiểu kháng kháng sinh nhất (4), với kiểu kháng kháng sinh chủ yếu là STR-NEO.
Bảng 1. Đặc điểm kháng kháng sinh của 5 loài Vibrio.
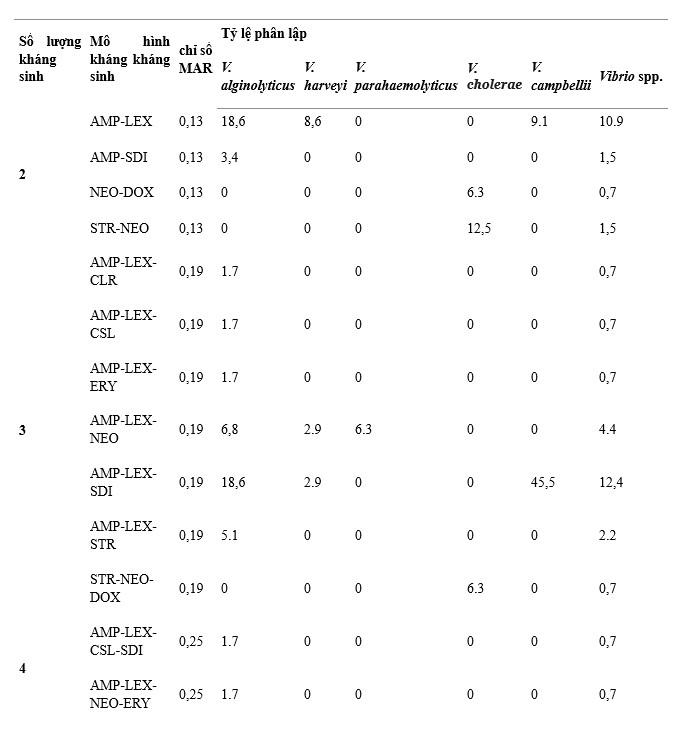
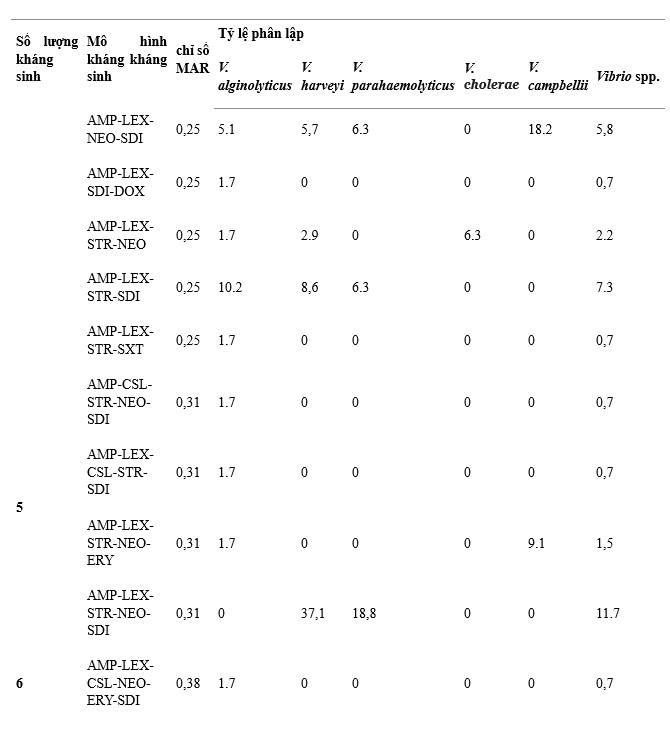

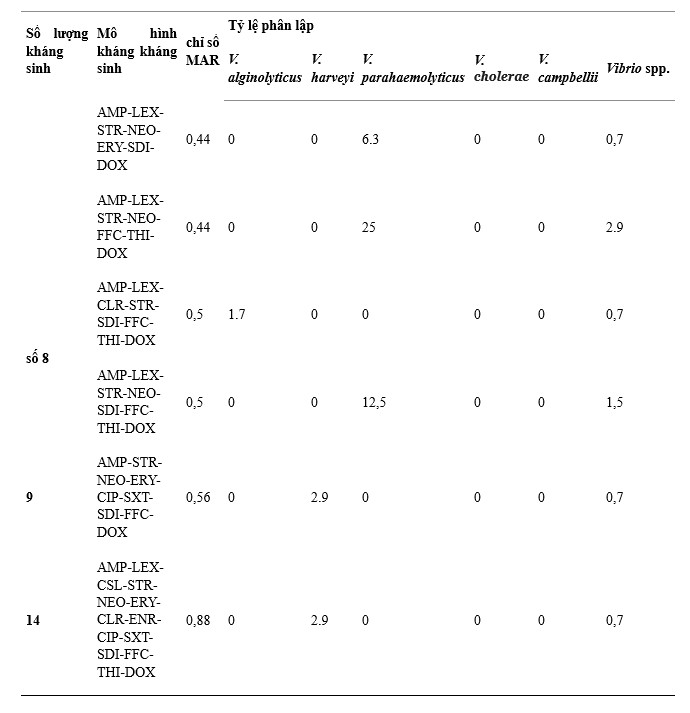
Chỉ số MAR dưới 137 chủng Vibrio phân lập là 0,13–0,88. Giá trị 0,13 MAR được phát hiện ở 13 phân lập V. alginolyticus, 3 phân lập V. harveyi, 3 phân lập V. cholerae và 1 phân lập V. campbellii, trong khi giá trị 0,88 MAR được quan sát thấy ở 1 phân lập V. harveyi có biểu hiện kháng thuốc 14/16 loại kháng sinh được thử nghiệm (Bảng 1). Chỉ số MAR giữa các loài lần lượt là 0,13–0,5 ở V. alginolyticus, 0,13–0,88 ở V. harveyi, 0,19–0,5 ở V. parahaemolyticus, 0,13–0,25 ở V. cholerae và 0,13–0,38 ở V. campbellii. Kết quả cho thấy 36,5% tổng số chủng phân lập có chỉ số MAR < 0,2, ghi nhận V. alginolyticus (n = 34, 24,8%), V. harveyi (n = 5, 3,6%), V. parahaemolyticus (n = 1)., 0,7%), V. cholerae (n = 4, 2,9%) và V. campbellii (n = 6, 4,4%) (Hình 4).
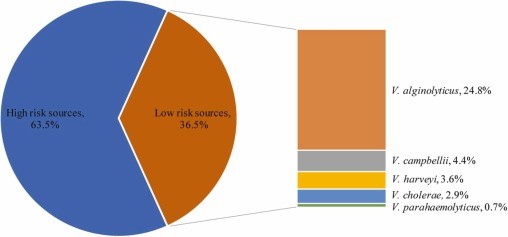
Hình 4. Phân loại chỉ số kháng đa kháng sinh (MAR) của các chủng vi khuẩn Vibrio.
ARPA là 0,28 trên 137 chủng Vibrio phân lập, trong đó V. parahaemolyticus (0,56) cho thấy ARPA cao nhất, tiếp theo là V. alginolyticus, V. harveyi và V. campbellii, với ARPA lần lượt là 0,44, 0,34 và 0,45 và V. cholerae cho ARPA thấp nhất là 0,25 (Hình 5).

Hình 5. Phân loại chỉ số phong phú mô hình kháng kháng sinh (ARPA) của 5 loài Vibrio.
3.5. Đánh giá rủi ro sinh thái của 5 loài Vibrio dựa trên RQ
RQ được tính toán của 5 loài Vibrio được trình bày trong Bảng 2. Tất cả các loài Vibrio đều có giá trị RQ thấp (<20) so với cefoperazone/sulbactam (0–15.3), enrofloxacin (0–2.9), ciprofloxacin (0–5.7), flumequine (0), flufenicol (0–6.3), tetracycline (0 –5.1) và doxycycline (0–14.3). V. alginolyticus, V. harveyi, V. parahaemolyticus và V. campbellii cho thấy RQ cao (>50) so với ampicillin (100), cefalexin (94,9–100) và neomycin (50–81,8), nhưng V. cholerae (0–6,3) hiển thị RQ thấp (<20). Ngược lại, V. alginolyticus, V. harveyi, V. cholerae và V. campbellii có RQ thấp (<20) so với sulfamethoxazole/trimethoprim (0–5,7) và thiamphenicol (0–3,4), trong khi V. parahaemolyticus (62,5) cho thấy cao RQ (>50).
Bảng 2. Chỉ số chỉ số kháng thuốc (RQs) của 5 chủng Vibrio phân lập ở các loài khác nhau.
3.6. Đánh giá gen kháng kháng sinh của các chủng Vibrio phân lập
Chỉ có 5 gen kháng thuộc aminoglycoside, quinolone, sulfonamid và integron được phát hiện dương tính với 137 chủng Vibrio phân lập (Hình bổ sung 3). Đối với β-lactam, macrolide, chloramphenicol và tetracyclines, không có gen kháng chọn lọc nào được phát hiện ở bất kỳ loài Vibrio nào. Đối với aminoglycoside, 2 gen kháng đã được phát hiện, 43,8% loài Vibrio chứa strA, 11,7% loài Vibrio có strB, trong khi 10,9% loài Vibrio chứa cả strA và strB (Bảng 3, Hình 6). Hơn nữa, 100% V. cholerae mang strA, 77,1% V. harveyi và 87,5% V. parahaemolyticus mang strA, và 68,8% V. parahaemolyticus cũng mang strB. Trong số các gen kháng quinolone được khảo nghiệm, chỉ có 2,9% chủng phân lập chứa QnrVC, và những chủng phân lập này là V. parahaemolyticus. Trong số các gen kháng sulfonamid được khảo nghiệm, chỉ có sul2 dương tính, chiếm 8,8% tổng số chủng phân lập được nghiên cứu. Chỉ có 1 chủng V. harveyi phân lập (2,9%) và 11 chủng V. parahaemolyticus (68,8%) biểu hiện sul2. Ngoài ra, chỉ Int4 (8,8%) được phát hiện trong số 4 loại integron và các phân lập dương tính được phát hiện ở 5,1% V. alginolyticus, 8,6% V. harveyi, 12,5% V. parahaemolyticus và 36,4% V. campbellii.
Bảng 3. Phân bố gen kháng kháng sinh của 5 loài Vibrio.
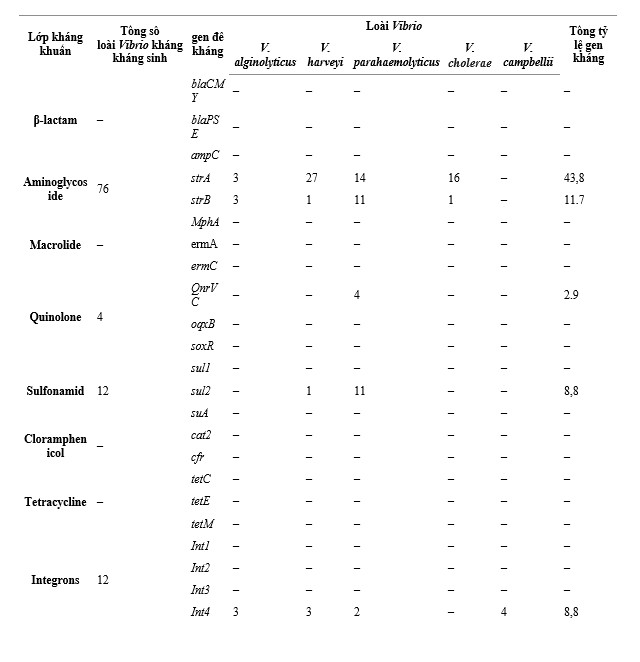
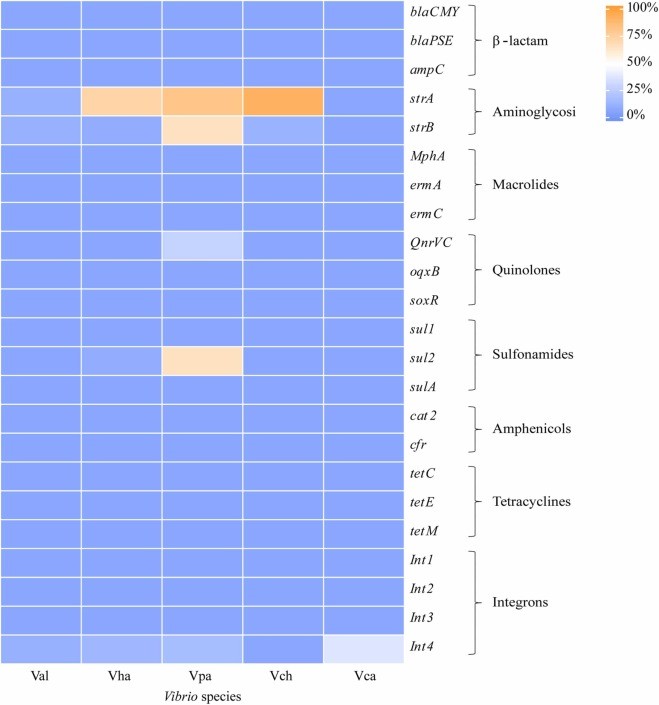
Hình 6. Tần số gen kháng kháng sinh giữa các loài Vibrio khác nhau. Val: V. alginolyticus; Vha: V. harveyi; Vpa: V. parahaemolyticus; Vch: V. cholerae; Vca: V. campbellii.
Phân tích của hiệp hội chỉ ra rằng các loài Vibrio có tính kháng cao (>50%) với ampicillin (89,1%) và cefalexin (86,1%), thuộc về β-lactam (Hình 3), nhưng các gen kháng β-lactam blaCMY, blaPSE, và ampC không được phát hiện (Hình 6). Tương tự, một số chủng Vibrio (2,2–34,3%) có khả năng kháng kháng sinh macrolide, chloramphenicol và tetracycline, trong khi không có chủng nào có gen kháng thuốc. Đối với các gen kháng dương tính, xét nghiệm chi bình phương cho thấy streptomycin tương quan với strA và strB (P < 0,05), và neomycin tương quan với strA (P < 0,05). Tuy nhiên, giá trị Cramer của chúng lần lượt là 0,28, 0,30 và 0,27, cho thấy mức độ liên kết thấp. Hơn nữa, QnrVC, sul2 và Int4 trước đây không tương quan với kiểu hình kháng kháng sinh của chúng (P > 0,05).
4. Thảo luận
Chi và loài mà vi khuẩn phân lập là một trong những yếu tố chính ảnh hưởng đến sự tương tác giữa độc lực và đặc tính kháng kháng sinh (Beceiro và cộng sự, 2013). Do đó, việc xác nhận các chủng Vibrio phân lập từ các vị trí mục tiêu trong nghiên cứu này đã đạt được dựa trên sự khuếch đại phân tử của rDNA ở thập niên 16, sau đó là các gen mục tiêu gyrB, toxR vh, toxR vp, ompW, hly đối với V. alginolyticus, V. harveyi, V. parahaemolyticus, V. cholerae và V. campbellii. Các nghiên cứu trước đây cũng đã sử dụng các phương pháp phân tử tương tự để xác nhận và phân định Vibrio phân lập thành loài (Guardiola-Avila và cộng sự, 2016, Okoh và cộng sự, 2015). Trong nghiên cứu này, việc xác nhận kiểu hình của Vibrio đã cho thấy 418 phân lập được cho là trong môi trường TCBS, trong khi xác nhận thêm bằng xét nghiệm rDNA vào thập niên 16 đã xác nhận rằng 312 phân lập thuộc chi Vibrio. V. alginolyticus, V. harveyi, V. parahaemolyticus, V. cholerae và V. campbellii là những loài chiếm ưu thế trong hệ thống nuôi tôm ở Trung Quốc. V. cholerae và V. parahaemolyticus có thể gây bệnh nghiêm trọng cho vật chủ là con người do ô nhiễm thực phẩm và nước (Gennari và cộng sự, 2012). V. alginolyticus, V. harveyi, V. parahaemolyticus và V. campbellii thuộc về các chủng dòng Harveyi, dòng mầm bệnh quan trọng nhất trong môi trường nước (Ruwandeepika và cộng sự, 2012, Gao và cộng sự, 2022). V. alginolyticus được phân lập thường xuyên nhất, tiếp theo là V. harveyi. Tương tự, Sadat và cộng sự (2021) phát hiện ra rằng V. alginolyticus có tỷ lệ phổ biến cao hơn ở động vật có vỏ.
Việc xác định các yếu tố độc lực là rất quan trọng để đánh giá đặc tính gây bệnh của vi khuẩn. Các sản phẩm vi sinh vật tiết ra có khả năng xâm nhập vào tế bào chủ và sử dụng cơ chế của chúng để hỗ trợ lây nhiễm (Kitamoto và cộng sự, 2016). Từ nghiên cứu này, 10 gen độc lực đã được phát hiện trong số các chủng Vibrio phân lập và tỷ lệ phát hiện là từ 11,4% đến 100%. Tương tự, De Silva và cộng sự (2019) đã phát hiện 8 gen độc lực ở Vibrio spp. phân lập từ sò điệp Yesso (Patinopecten Yessoensis) được bán trên thị trường ở Hàn Quốc, với tỷ lệ nhiễm từ 3% đến 100%. Tuy nhiên, Mohamad và cộng sự. (2019) phát hiện Vibrio spp. phân lập từ cá biển nuôi lồng ở Malaysia có tỷ lệ phát hiện gen độc cao (60–100%). So với kết quả đó, Vibrio spp. trong hệ thống nhân giống P. vannamei ở Trung Quốc có nguy cơ tiềm tàng mang và truyền các gen độc lực. Trong số các gen độc lực của V. parahaemolyticus được thử nghiệm, chỉ có 1 gen độc lực (tlh) dương tính, cho thấy V. parahaemolyticus có độc lực cao không phổ biến trong hệ thống nuôi tôm ở Trung Quốc. Tuy nhiên, Khouadja và cộng sự. (2014) đã tìm thấy 6 gen độc lực ở V. parahaemolyticus phân lập từ nước thải đã qua xử lý, tỷ lệ phát hiện có thể lên tới 100%. Hiện tượng này có thể là do việc phát hiện và ngăn chặn hiệu quả các vi khuẩn gây bệnh tiềm ẩn trong hệ thống nuôi tôm ở Trung Quốc, đồng thời làm giảm đáng kể sự lây lan của các gen độc lực trong môi trường nước. So với các loài Vibrio khác được thử nghiệm, cả 5 gen độc lực của V. campbellii trong nghiên cứu này đều được phát hiện dương tính và hầu hết chúng đều có tỷ lệ phát hiện rất cao. Tương tự, Mohamad và cộng sự (2019) đã phát hiện 5 gen độc lực ở V. campbellii phân lập từ cá biển nuôi lồng ở Malaysia với tỷ lệ phát hiện cao và Kumar và cộng sự (2021) cũng phát hiện 11 gen độc lực ở V. campbellii từ các trại giống tôm Ấn Độ với tỷ lệ phát hiện là 100%. Có thể thấy V. campbellii mang gen độc có thể lan rộng trong môi trường nước, hiện tượng này cần được quan tâm nghiêm túc.
Đánh giá đặc tính kháng kháng sinh của mầm bệnh và nguy cơ sinh thái tiềm ẩn của nó là mối quan tâm lớn trên toàn thế giới đối với nghiên cứu và phát triển y học (Xie và cộng sự, 2017). Kết quả cho thấy Vibrio phân lập trong hệ thống nuôi tôm có khả năng kháng cao với ampicillin (89,1%) và cehalexin (86,1%), cả hai đều là kháng sinh thế hệ đầu tiên. Các kháng sinh thế hệ đầu tiên bao gồm ampicillin được cho là tác nhân chính gây ra tình trạng kháng thuốc, do đó làm giảm tính nhạy cảm và do đó làm giảm hiệu quả ức chế Vibrio của loại thuốc này (Sudha và cộng sự, 2014). Các loài Vibrio được xác định trong thí nghiệm này nhạy cảm (<10%) với flumequine, cefoperazone/sulbactam, clarithromycin, enrofloxacin, ciprofloxacin, sulfamethoxazole/trimethoprim, flufenicol, thiamphenicol, tetracycline và doxycycline. Và nhiều chủng Vibrio cho thấy độ nhạy kháng sinh trung bình với streptomycin (42,3%), neomycin (39,4%) và erythromycin (64,2%).
Phân tích chỉ số đặc tính kháng kháng sinh cho thấy 63,5% tổng số chủng phân lập có chỉ số MAR > 0,2. Mohamad và cộng sự. (2019) cho thấy khoảng 75% chủng Vibrio phân lập có chỉ số MAR > 0,2 trên cá bị bệnh ở Malaysia. Nhưng điều này đối với các đợt bùng phát dịch bệnh tự nhiên ở cá biển/cửa sông ở Ấn Độ cho thấy 38,89% số chủng Vibrio phân lập có MAR> 0,20 (Sony và cộng sự, 2021). Kết quả cho thấy chủng MAR Vibrio có tỷ lệ lưu hành cao trong hệ thống nhân giống P. vannamei ở Trung Quốc, điều này cần được xem xét nghiêm túc. ARPA của Vibrio spp. bị cô lập bởi Đặng và cộng sự (2020) từ cá biển mắc bệnh ở Nam Trung Quốc là 0,54, ngược lại, ARPA của Vibrio spp. trong nghiên cứu này là 0,28, cho thấy Vibrio spp. trong nghiên cứu này có sự biến đổi thấp về kiểu hình kháng thuốc kháng sinh, điều này càng chỉ ra rằng kiểu hình trao đổi chất của các loài Vibrio trong hệ thống nuôi tôm ở Trung Quốc là nhất quán và dễ điều chỉnh. Tuy nhiên, V. parahaemolyticus ghi nhận ARPA cao là 0,56, trong khi Adesiyan và cộng sự (2021) phát hiện ra rằng V. parahaemolyticus thu hồi từ các con sông ở Tây Nam Nigeria có mức độ đề kháng thấp là 0,19. Vì vậy, chỉ số ARPA của loài Vibrio có thể liên quan đến môi trường cách ly của loài Vibrio. RQ thấp (<20) của 5 loài Vibrio chống lại cefoperazone/sulbactam, enrofloxacin, ciprofloxacin, flumequine, flufenicol, tetracycline và doxycycline, và có thể hàm ý không có hoặc áp lực chọn lọc thấp đối với khả năng kháng các kháng sinh này trong môi trường nuôi tôm được lấy mẫu.
Các gen kháng kháng sinh có thể được phổ biến giữa các vi khuẩn khác nhau thông qua quá trình biến đổi, liên hợp hoặc tải nạp (Manjusha và Sarita, 2011). Ngoài ra, các gen kháng kháng sinh, đặc biệt là các gen liên quan đến yếu tố di truyền di động, có thể truyền sang các mầm bệnh vi khuẩn ảnh hưởng đến con người (Shakerian và cộng sự, 2017). Việc phát hiện nhiều loại và tỷ lệ phát hiện gen kháng kháng sinh cao cho thấy nguy cơ truyền gen kháng kháng sinh cao, điều này cũng thể hiện nguy cơ cao vi khuẩn tạo ra tình trạng kháng thuốc. Trong nghiên cứu này chỉ phát hiện được 4 trong 8 loại gen kháng kháng sinh, chỉ có 5 loại gen kháng, trong đó strB (11,7%), QnrVC (2,9%), sul2 (8,8%) và Int4 (8,8%) đều có tỷ lệ phát hiện thấp ngoại trừ strA (43,8%), cho thấy nguy cơ truyền gen kháng thấp trong hệ thống nhân giống P. vannamei ở Trung Quốc. Phân tích tương quan giữa kiểu hình kháng kháng sinh và các gen kháng kháng sinh bằng xét nghiệm chi bình phương cho thấy streptomycin có mối tương quan dương tính thấp với strA (giá trị của Cramer là 0,28; P < 0,05) và strB (giá trị của Cramer là 0,30; P < 0,05), và neomycin là cũng có tương quan dương thấp với strA (giá trị của Cramer là 0,27; P < 0,05). Sony và cộng sự. (2021) đã tìm thấy mối tương quan dương đáng kể giữa tình trạng kháng oxytetracycline và tetB khi sử dụng cùng một phương pháp. Các gen kháng kháng sinh β-lactam blaCMY, blaPSE và ampC không được phát hiện ở bất kỳ chủng Vibrio nào, trong khi tất cả các loài Vibrio đều có khả năng kháng cao với kháng sinh β-lactam ampicillin và ceshalexin. Sự khác biệt như vậy giữa kiểu hình kháng kháng sinh và tỷ lệ phổ biến của gen kháng kháng sinh đã được nhiều tác giả quan sát (Đặng và cộng sự, 2006, Raissy và cộng sự, 2012, Yu và cộng sự, 2022), có thể là do kiểu hình kháng khuẩn có thể gây ra được biểu hiện dưới sự kích thích của nhiều yếu tố di truyền khác nhau (Van và cộng sự, 2020). Một cách giải thích khác có thể là kiểu hình kháng khuẩn không chỉ được điều hòa bởi các gen kháng thuốc mà còn bởi cấu trúc màng và chuyển hóa sinh lý.
5. Kết luận
Sự phân bố của các loài Vibrio và nguy cơ độc lực và khả năng kháng kháng sinh của các mầm bệnh chính trong hệ thống nuôi tôm là nguy cơ lây truyền bệnh và rủi ro sinh thái tiềm ẩn trong hệ thống thủy sản. Trên cơ sở đó, sự phân bố quần thể Vibrio, độc lực, tính kháng kháng sinh và nguy cơ sinh thái của các chủng Vibrio chiếm ưu thế trong nuôi tôm bố mẹ, chương trình nhân giống, môi trường nuôi và các yếu tố đầu vào ngoại sinh (mồi sinh học, v.v.) trong hệ thống nhân giống Penaeus vannamei ở phía Nam. Trung Quốc được đánh giá kết quả cho thấy V. alginolyticus, V. harveyi, V. parahaemolyticus, V. cholerae và V. campbellii là những loài chiếm ưu thế trong chương trình nhân giống tôm của Trung Quốc. Các chủng vi khuẩn Vibrio phân lập trong hệ thống nuôi tôm có nguy cơ tiềm tàng mang và truyền các gen độc lực. Và phân tích chỉ số đặc tính kháng kháng sinh chỉ ra rằng kiểu hình trao đổi chất của loài Vibrio trong hệ thống nuôi tôm ở Trung Quốc có hiệu ứng phù hợp và áp lực chọn lọc thấp. Hơn nữa, kết quả phát hiện gen kháng kháng sinh cho thấy nguy cơ lây truyền gen kháng kháng sinh trong hệ thống nuôi tôm là thấp. Các kết quả nghiên cứu có liên quan cung cấp cơ sở lý thuyết để hiểu rõ nguy cơ mắc các bệnh Vibrio chính và rủi ro sinh thái trong hệ thống nuôi trồng thủy sản ở Trung Quốc, đồng thời cung cấp hỗ trợ dữ liệu chung cho sự phát triển lành mạnh và bền vững của ngành thủy sản.
Theo Yongxiang Yu, Miaomiao Tang, Yingeng Wang, Meijie Liao, Chunyuan Wang, Xiaojun Rong, Bin Li, Jianlong Ge, Yingli Gao, Xuan Dong, Zheng Zhang
Nguồn: https://www.sciencedirect.com/science/article/pii/S0147651323001197
Biên dịch: Nguyễn Thị Quyên – Bình Minh Capital
Xem thêm:
- Probiotic Bacillus Mang Lại Lợi Ích Trong Nuôi Cá Rô Phi Tại Brazil Ở Điều Kiện Đầy Thách Thức
- Ảnh Hưởng Của Probiotic Loài Bacillus (Bacillus subtilis Và B. licheniformis) Đến Chất Lượng Nước, Phản Ứng Miễn Dịch Và Sức Đề Kháng Của Tôm Chân Trắng (Litopenaeus vannamei) Chống Lại Nhiễm Nấm Fusarium solani
- Probiotic Trong Nuôi Trồng Thủy Sản