
 English
English
Kỹ Thuật Nuôi, Tin tức
Phần 2: Ảnh Hưởng Của Điều Kiện Ánh Sáng Đến Hiệu Suất Tăng Trưởng Và Thành Phần Sinh Hóa Của Cua Bùn Cái Sau Khi Giao Phối (Scylla paramamosain)
3. Kết quả
3.1. Các biến số sinh trưởng và sinh sản
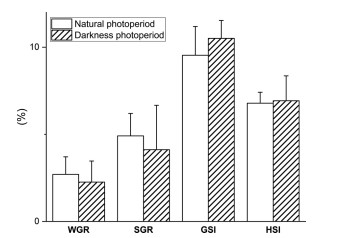
WGR, SGR, GSI và HSI của S. paramamosain nuôi cấy trong các điều kiện ánh sáng khác nhau được thể hiện trong Hình 1. WGR, SGR, HSI và GSI không bị ảnh hưởng bởi điều kiện ánh sáng (P > 0,05). Tuy nhiên, GSI của S. paramamosain cái trong điều kiện bóng tối hoàn toàn cao hơn một chút so với GSI trong điều kiện ánh sáng bình thường (mặc dù không quan sát thấy sự khác biệt về mặt thống kê; P > 0,05).
3.2. Thành phần cơ thể
Bảng 1 cho thấy các thành phần (độ ẩm, tro, protein thô và lipid thô) trong cơ, gan tụy và buồng trứng. S. paramamosain cái được nuôi cấy trong điều kiện bóng tối hoàn toàn có hàm lượng tro cao hơn trong cơ (23,27% khối lượng khô) và buồng trứng (7,56% khối lượng khô) so với những con trong điều kiện ánh sáng bình thường. Tuy nhiên, hàm lượng protein trong cơ (85,26% khối lượng khô) và buồng trứng (69,71% khối lượng khô) thấp hơn so với hàm lượng protein trong điều kiện ánh sáng bình thường (cơ, 88,08% khối lượng khô; buồng trứng, 72,14% khối lượng khô) (P < 0,05).
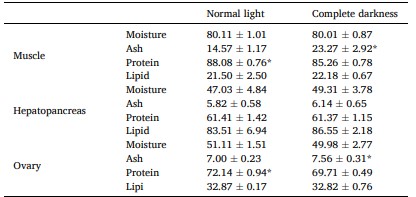
Sự khác biệt đáng kể được chỉ ra bằng dấu * (P < 0,05). Các giá trị là trung bình (± SD).
3.3. Hàm lượng carotenoid
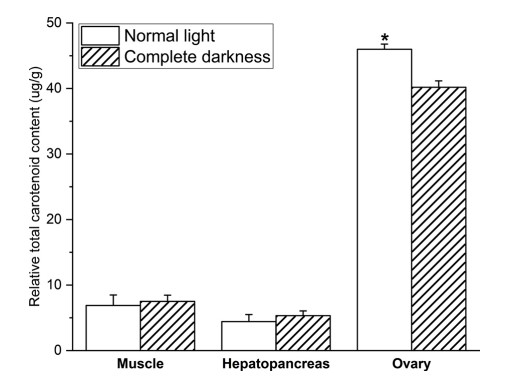
RTCC trong các mô khác nhau bao gồm cơ, gan tụy và buồng trứng được thể hiện trong Hình 2. RTCC trong buồng trứng của cua được nuôi trong bóng tối hoàn toàn (40,19 ug/g) thấp hơn đáng kể so với RTCC trong điều kiện ánh sáng bình thường (45,98 ug/g) (P < 0,05).
3.4. Hàm lượng axit amin
Hàm lượng axit amin của cơ được thể hiện trong Bảng 2. Cơ của cua được nuôi trong bóng tối hoàn toàn có hàm lượng axit aspartic, threonine, glycine, methionine, leucine, tyrosine, lysine, arginine, tổng axit amin (TAA), axit amin hương vị (FAA) và axit amin thiết yếu (EAA) thấp hơn đáng kể (P < 0,05).
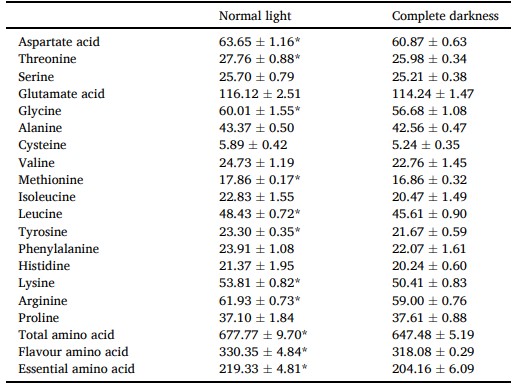
Axit amin hương vị bao gồm: Axit aspartic, Axit glutamic, Glycine, Alanine, Methionine, Phenylalanine; Axit amin thiết yếu bao gồm: Threonine, Valine, Methionine, Isoleucine, Leucine, Phenylalanine, Lysine. Sự khác biệt đáng kể được chỉ ra bằng dấu * (P < 0,05). Giá trị là trung bình (± SD).
Bảng 3 trình bày hàm lượng axit amin của gan tụy. Hàm lượng glycine (8,25 mg/g) và isoleucine (4,75 mg/g) trong gan tụy của cua được nuôi trong bóng tối hoàn toàn cao hơn đáng kể so với hàm lượng trong điều kiện ánh sáng bình thường (P < 0,05).
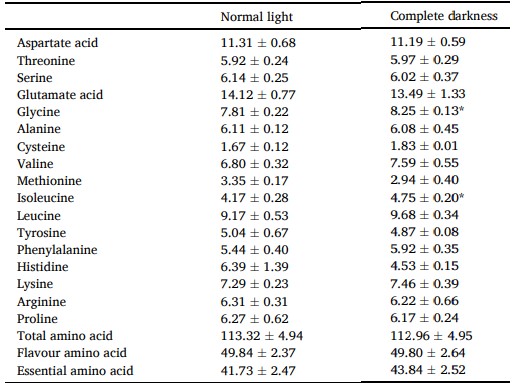
Axit amin hương vị bao gồm: Axit aspartic, Axit glutamic, Glycine, Alanine, Methionine, Phenylalanine; Axit amin thiết yếu bao gồm: Threonine, Valine, Methionine, Isoleucine, Leucine, Phenylalanine, Lysine. Sự khác biệt đáng kể được chỉ ra bằng dấu * (P < 0,05). Giá trị là trung bình (± SD).
Hàm lượng axit amin của buồng trứng được thể hiện trong Bảng 4. Hàm lượng proline trong buồng trứng của cua được nuôi trong bóng tối hoàn toàn thấp hơn so với nhóm điều kiện ánh sáng bình thường (P < 0,05).
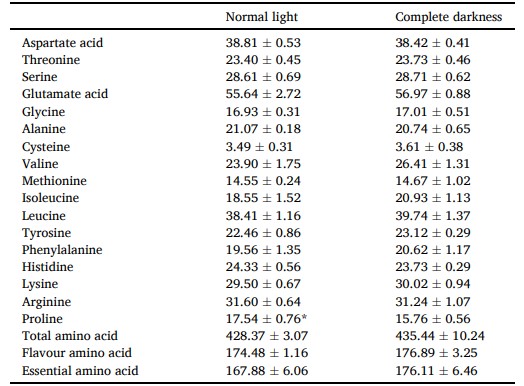
Axit amin hương vị bao gồm: Axit aspartic, Axit glutamic, Glycine, Alanine, Methionine, Phenylalanine; Axit amin thiết yếu bao gồm: Threonine, Valine, Methionine, Isoleucine, Leucine, Phenylalanine, Lysine. Sự khác biệt đáng kể được chỉ ra bằng dấu * (P < 0,05). Các giá trị là trung bình (± SD).
3.5. Hàm lượng axit béo
Hàm lượng axit béo của gan tụy được thể hiện trong Bảng 5. Hàm lượng PUFA 20:1, 20:2n-6, và n-6 trong gan tụy của cua được nuôi trong bóng tối hoàn toàn cao hơn đáng kể so với điều kiện ánh sáng bình thường (P < 0,05). Ngược lại, gan tụy của cua trong bóng tối hoàn toàn có tỷ lệ 18:1 và n-3/n-6 thấp hơn đáng kể so với gan tụy trong điều kiện ánh sáng bình thường (P < 0,05).
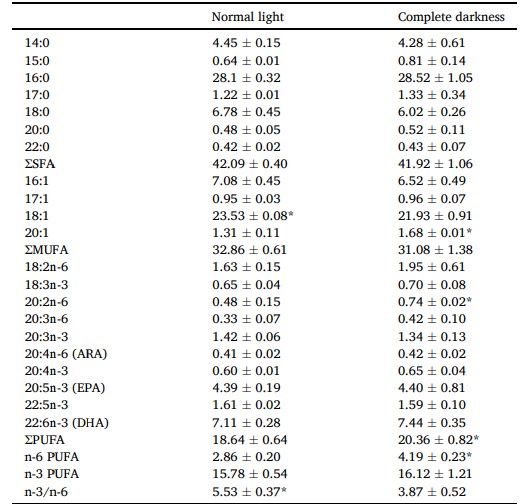
Sự khác biệt đáng kể được biểu thị bằng dấu * (P < 0,05). Giá trị là trung bình (± SD).
Buồng trứng của cua được nuôi trong bóng tối hoàn toàn có hàm lượng 20:0, 22:0, 20:1, 18:2n-6, 20:3n-6 và n-6 cao hơn, nhưng tỷ lệ 20:3n-3, 22:5n-3 và n-3/n-6 thấp hơn so với gan tụy được nuôi trong điều kiện ánh sáng bình thường (P < 0,05; Bảng 6).
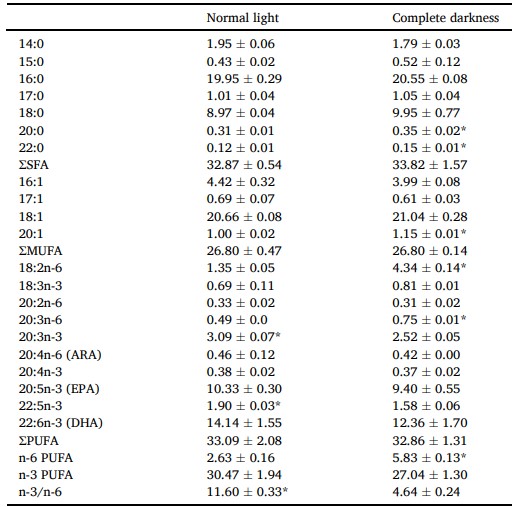
Sự khác biệt đáng kể được chỉ ra bằng dấu * (P < 0,05). Các giá trị là trung bình (± SD).
3.6. Đặc điểm hình thái
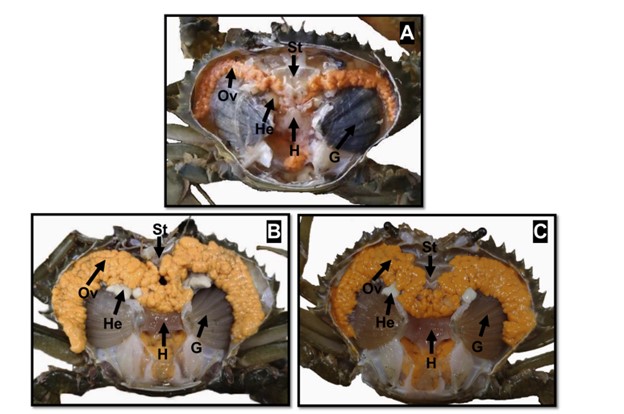
Hình thái bên trong của con cái S. paramamosain dưới hai điều kiện ánh sáng được trình bày trong Hình 3. Thí nghiệm ảnh hưởng đến màu sắc và kích thước của buồng trứng. Khi bắt đầu thí nghiệm, buồng trứng ở giai đoạn IV (giai đoạn tiền trưởng thành) và có kích thước nhỏ (Hình 3A). Sau 1 tháng thí nghiệm, buồng trứng mở rộng, lớn hơn và đạt đến giai đoạn V (giai đoạn trưởng thành) (Hình 3B, C). Buồng trứng của cua được nuôi trong điều kiện ánh sáng bình thường có màu cam sáng (có thể là do hàm lượng carotenoid cao hơn), trong khi những con ở trong bóng tối hoàn toàn có màu vàng nhạt. Các giai đoạn phát triển của buồng trứng được xác định dựa trên kích thước, màu sắc và hình thái theo Q. Wu và cộng sự (2020).
4. Thảo luận
4.1. Hiệu suất tăng trưởng
Chúng tôi đã nghiên cứu WGR, SGR và HSI ở S. paramamosain cái được nuôi trong bóng tối hoàn toàn (0 ± 0 lx; 0 giờ sáng L: 24 giờ tối) và điều kiện ánh sáng bình thường (7,00 ± 1,50 lx; 12 giờ sáng: 12 giờ tối). WGR và SGR biểu thị hiệu suất tăng trưởng và nuôi cấy. HSI được coi là một chỉ số quan trọng về trạng thái dự trữ năng lượng trong gan tụy (Wu và cộng sự, 2020). Một số nghiên cứu cho thấy cường độ ánh sáng và quang kỳ ảnh hưởng đến hiệu suất tăng trưởng của giáp xác. Hầu hết các nghiên cứu đều cho thấy ấu trùng và ấu trùng giáp xác có hiệu suất nuôi cấy tốt hơn trong quang pha dài với cường độ ánh sáng cao hơn. Ví dụ, bóng tối hoàn toàn dẫn đến tỷ lệ sống và tốc độ phát triển của ấu trùng thấp hơn, trong khi quang kỳ 18 L: 6D là chế độ quang kỳ phù hợp nhất để nuôi ấu trùng P. pelagicus (Andres và cộng sự, 2010). Ấu trùng của J. edwardsii có tốc độ tăng trưởng nhanh hơn khi tăng chiều dài ngày (Bermudes và Ritar, 2008). Tương tự như vậy, ấu trùng P. gigas có thời gian lột xác ngắn hơn và phát triển nhanh hơn dưới cường độ ánh sáng cao (Gardner và Maguire, 1998). Không giống như ấu trùng,rất ít thông tin có sẵn về tác động của cường độ ánh sáng và quang kỳ đến hiệu suất tăng trưởng của giáp xác trưởng thành. Trong nghiên cứu này, không có sự khác biệt đáng kể nào giữa hai nhóm về hiệu suất tăng trưởng (WGR và SGR). Những kết quả này có thể là do kích thước lớn hơn của cua trưởng thành. Ở giáp xác, để phát triển, lớp vỏ cũ phải thường xuyên được thay thế bằng lớp vỏ ngoài mới lớn hơn. Vì giai đoạn lột xác dài hơn khi cá thể lớn lên nên cua lớn phát triển chậm hơn cua nhỏ (Farhadi và Jensen, 2016; Tao và cộng sự, 2014).
4.2. Hiệu suất sinh sản
GSI cung cấp thông tin liên quan đến sự trưởng thành về mặt sinh dục và trạng thái phát triển của tinh hoàn hoặc buồng trứng (Flores và cộng sự, 2015). Trong nghiên cứu này, bóng tối hoàn toàn làm tăng nhẹ GSI (P > 0,05) của S. paramamosain cái khi so sánh với chế độ quang kỳ tự nhiên. Bóng tối hoàn toàn tác đông tích cực đến tốc độ sinh sản đã được báo cáo ở một số loài giáp xác bao gồm P. leptodactylus (Harlıoglu và Duran, 2010), C. sapidus (Bembe và cộng sự, 2017), P. llamasi (Carmona-Osalde và cộng sự, 2002) và P. merguiensis (Hoang và cộng sự, 2002a, 2002b,2002c). Ví dụ, C. sapidus cái sinh sản nhiều nhất trong điều kiện bóng tối liên tục (Bembe và cộng sự, 2017). Nuôi P. leptodactylus cái trong bóng tối hoàn toàn thúc đẩy sinh sản và tăng tỷ lệ con cái đẻ trứng (Harlıoglu và Duran, 2010). Giải thích cho tỷ lệ sinh sản cao hơn trong bóng tối hoàn toàn là bóng tối có thể có tác dụng ức chế cơ quan X bằng cách giảm kích thích cơ quan này (Farhadi và Harlıoglu, 2019b). Cơ quan X là trung tâm điều hòa nội tiết thần kinh chính và nằm ở cuống mắt của giáp xác (Fingerman, 1987). Cơ quan X điều chỉnh sinh sản của giáp xác bằng cách tiết hormone ức chế lột xác (MIH), hormone ức chế tuyến sinh dục/sinh noãn hoàng (GIH hoặc VIH) và CHH (hormone tăng đường huyết ở giáp xác) (Bocking và cộng sự, 2002). Kích thích cơ quan X làm tăng tiết các hormone thần kinh này và ức chế quá trình lột xác và sinh sản (Bocking và cộng sự, 2002; Lacombe và cộng sự, 1999). Việc cắt bỏ cuống mắt làm tăng tốc độ trưởng thành của tuyến sinh dục và làm tăng tỷ lệ sinh sản (Fingerman, 1987).
4.3. Hàm lượng protein và axit amin
Quá trình chuyển hóa protein và axit amin ở giáp xác dưới các yếu tố môi trường khác nhau như cường độ ánh sáng và quang kỳ vẫn chưa rõ ràng (Farhadi và Harlıoglu, 2019b). Trong nghiên cứu này, hàm lượng protein và axit amin trong cơ và hàm lượng protein trong buồng trứng giảm trong điều kiện bóng tối hoàn toàn. Những kết quả này trái ngược với những phát hiện trước đây trong đó việc cắt bỏ cuống mắt làm tăng quá trình dự trữ protein, carbohydrate, lipid và ức chế quá trình tiêu thụ chất hữu cơ (Murugesan và cộng sự, 2008; Sun và cộng sự, 2014). Do đó, có thể cho rằng con đường lắng đọng protein và axit amin trong bóng tối hoàn toàn có thể không giống như trong cơ chế cắt bỏ cuống mắt. Protein là yếu tố quan trọng cho quá trình thụ tinh và phát triển phôi ở cua bùn (Waiho và cộng sự, 2018). Do đó, việc giảm protein trong buồng trứng trong bóng tối hoàn toàn có thể làm suy giảm chất lượng ấu trùng.
FAA đóng vai trò quan trọng trong việc hình thành hương vị thịt. Ví dụ, axit aspartic và axit glutamic có vị mặn (hay còn gọi là umami) và tạo ra vị umami. Vị umami của thịt cua có sự đóng góp đáng kể của glutamate. Arginine, phenylalanine và tyrosine tạo ra vị đắng và chúng có nhiều trong các loại hải sản như cua (Shao và cộng sự, 2013). Phenylalanine và tyrosine cũng là những thành phần chính tạo nên vị mặn. Glycine và alanine tạo ra vị ngọt dễ chịu (Sarower và cộng sự, 2012). Arginine, glycine, alanine và axit glutamic có tác động mạnh đến hương vị thịt cua (Chen và Zhang, 2007). S. paramamosain cái được nuôi trong điều kiện ánh sáng bình thường có hàm lượng FAA cao hơn trong cơ. Do đó, có thể kết luận rằng nuôi S. paramamosain trong điều kiện ánh sáng bình thường có thể mang lại hương vị ngon hơn và giá trị dinh dưỡng cao hơn so với nuôi trong bóng tối hoàn toàn.
4.4. Hàm lượng lipid và axit béo
Lipid buồng trứng, SFA, MUFA và PUFA đóng vai trò quan trọng trong quá trình sinh sản như trưởng thành buồng trứng, là nguồn dự trữ năng lượng cho quá trình sản xuất trứng trong tương lai, phát triển phôi và tăng trưởng ấu trùng (Alava và cộng sự, 2007a; Wu và cộng sự, 2020). Gan tụy là cơ quan quan trọng nhất cung cấp chất nền và chuyển hóa năng lượng thành các dạng có thể sử dụng trong quá trình sinh sản (Kyomo, 1988). Axit béo là thành phần quan trọng của cơ thể con người và cần có thành phần cân bằng các axit béo thiết yếu để duy trì sức khỏe của con người. Ví dụ, các axit béo thiết yếu như PUFA rất quan trọng cho sự phát triển của hệ thần kinh trung ương ở phôi người (Muskiet và cộng sự, 2006). Các chuyên gia dinh dưỡng khuyến cáo rằng tỷ lệ PUFA n-3/n-6 trong chế độ ăn uống phải đạt ít nhất 0,1− 0,2 (FAO, 2008). Tỷ lệ n-3/n-6 trong gan tụy và buồng trứng ở cả hai nhóm đều cao, do đó, chúng có thể được định nghĩa là các loại hải sản lành mạnh giàu n-3 PUFA (FAO, 2008). Tuy nhiên, buồng trứng và gan tụy của cua trong điều kiện ánh sáng bình thường cho thấy tỷ lệ n-3/n-6 cao hơn. Ngược lại, MUFA gan tụy, PUFA n-6 gan tụy và buồng trứng trong bóng tối hoàn toàn cao hơn so với điều kiện ánh sáng bình thường.
4.5. Hàm lượng carotenoid
Giống như hầu hết các loài động vật, giáp xác không thể tổng hợp carotenoid và phải lấy chúng từ chế độ ăn (Wade và cộng sự, 2017). Carotenoid đóng vai trò quan trọng trong việc tạo màu cho buồng trứng từ vàng nhạt đến đỏ cam sáng trong quá trình phát triển buồng trứng (Ghazali và cộng sự, 2017). Carotenoid cũng đóng vai trò là chất chống oxy hóa và đóng vai trò quan trọng trong việc bảo vệ trứng và phát triển mắt ở ấu trùng giáp xác (Dall và cộng sự, 1995; Tantikitti và cộng sự, 2015; Wu và cộng sự, 2020). Carotenoid buồng trứng gia tăng trong quá trình trưởng thành của buồng trứng chỉ ra sự tham gia có thể có của carotenoid vào quá trình phát triển trứng ở S. paramamosain (Wu và cộng sự, 2020). Nghiên cứu của chúng tôi cho thấy bóng tối hoàn toàn làm giảm sự tích tụ carotenoid trong buồng trứng của S. paramamosain. Penaeus vannamei được nuôi trong điều kiện cường độ ánh sáng cao (2500 lx) cho thấy astaxanthin tích tụ hơn so với những con được nuôi trong điều kiện bóng tối hoàn toàn (You và cộng sự, 2006). Nguyên nhân dẫn đến sự tích tụ carotenoid thấp ở giáp xác trong điều kiện bóng tối hoàn toàn vẫn chưa được biết rõ. Có rất ít thông tin về các cơ chế phân tử và di truyền cơ bản cho quá trình hấp thụ, vận chuyển và tích tụ mô của carotenoid ở bất kỳ loài động vật nào (Wade và cộng sự, 2017).
5. Kết luận
Tóm lại, nghiên cứu này đã đánh giá các chỉ tiêu WGR, SGR, HSI, GSI, cùng với thành phần và hàm lượng carotenoid, axit béo và axit amin của cua cái Scylla paramamosain sau giao phối khi được nuôi trong hai điều kiện chiếu sáng khác nhau: bóng tối hoàn toàn (0 ± 0 lx; 0 giờ sáng : 24 giờ tối) và ánh sáng bình thường (7,00 ± 1,50 lx; 12 giờ sáng : 12 giờ tối). Kết quả cho thấy điều kiện bóng tối hoàn toàn có ảnh hưởng đáng kể đến thành phần sinh hóa của cua cái S. paramamosain. Cụ thể, các cá thể được nuôi trong bóng tối hoàn toàn có hàm lượng axit amin và protein trong cơ thấp hơn so với nhóm nuôi trong điều kiện ánh sáng bình thường. Đồng thời, bóng tối hoàn toàn cũng làm suy giảm hàm lượng carotenoid và protein trong buồng trứng. Sự suy giảm carotenoid và protein ở buồng trứng dưới điều kiện bóng tối hoàn toàn có thể là những yếu tố hạn chế khả năng sinh sản của cua cái S. paramamosain. Do đó, các nghiên cứu tiếp theo cần tập trung đánh giá hiệu quả sinh sản của cua cái nuôi trong điều kiện này khi được cung cấp khẩu phần có hàm lượng carotenoid và protein cao.
Theo Ardavan Farhadi, Zhi Huang, Bixun Qiu, Mhd Ikhwanuddin, Hongyu Ma
Nguồn: https://www.sciencedirect.com/science/article/pii/S2352513421002234
Biên dịch: Nguyễn Thị Quyên – Bình Minh Capital
Xem thêm:
- Phân Tích Bệnh Phân Trắng (WFD) do Vibrio sp. Và Dinoflagellata Ở Tôm Thẻ Chân Trắng (Litopenaeus vannamei) Ở Ao Nuôi Nước Lợ
- Đâu Là Giải Pháp Thay Thế Khả Thi Cho Việc Cắt Bỏ Cuống Mắt Tôm?
- Ảnh Hưởng Của Việc Cho Ăn Trong Quá Trình Loại Bỏ Mùi Vị Khó Chịu Đến Sự Bài Tiết Geosmin Ở Cá Rô Phi